Magnesium-Bulletin 3 (Suppl 1a)/1981
Magnesium Requirements in Human Nutrition
By M. S. Seelig
Department of Medicine, Goldwater Memorial Hospital, New York
University Medical Center, New York, USA
Go to figures for this article, Magnesium Requirements in Human
Nutrition(figures)
German Summary
English Summary
French Summary
Introduction and article in English
Zusammenfassung
Der Bedarf an Magnesium wird unterschätzt. Dies hat die
folgenden Gründe: (1) Es besteht die falsche Auffassung,
daß der tägliche Bedarf die Menge ist, die Zeichen und
Symptome schweren Mg-Mangels oder Hypomagnesämie vermeidet;
(2) Schwierigkeiten bei der Bestimmung des zellulären Mg;
Plasma-Mg ist ein schlechter Indikator für den Mg-Status;
(3) Die Magnesium-Bilanz wird aufrechterhalten bei suboptimalen
Aufnahmen und Gewebekonzentrationen; (4) Der Bedarf ist
gesteigert im Wachstum, während Reparaturvorgängen,
Streß, Diätimbalanzen und durch Umweltfaktoren.
Mg-Mangel verursacht viele Anomilitäten; sogar parenteral
appliziertes Mg ist im allgemeinen sicher.
Schlußfolgerung: Bevor keine endgültigen Daten
vorliegen über die optimale Aufnahme unter physiologischen
und pathophysiologischen Bedingungen, sollte die Mg-Aufnahme
für junge Erwachsene auf 6 bis 10 mg/kg/Tag erhöht
werden und um den doppelten Betrag bei aktiven Anabolismus
und/oder unter Streßbedingungen.
Summary
Magnesium requirements are underestimated. This is due to: (1)
the misconception that the daily requirement is the amount that
prevents signs and symptoms of severe deficiency or
hypomagnesemia; (2) the difficulties in assaying cellular Mg,
plasma Mg being a poor index of body status; (3) the maintenance
of Mg-balance at suboptimal intakes and tissue levels; and (4)
the increased needs caused by growth, development, repair,
stress, dietary imbalances, and environmental factors. Mg
deficiency causes many abnormalities; even parenteral Mg is
generally safe, Thus, until definitive data are available as to
optimal intakes under different physiologic and pathologic
conditions, Mg-intakes should be increased to 6-10 mg/kg/day for
young adults, and to twice that much for those undergoing active
anabolism, or under stress.
Résumé
Les besoins en Mg sont sous-estimés. Ceci est
dû:
1) à la conception erronée selon laquelle le
besoin quotidien est la quantité qui prévient les
signes et les symptomes du déficit magnésique grave
et de l'hypomagnésémie,
2) aux difficultés dans l'évaluation du Mg
cellulaire, le Mg plasmatique étant un mauvais indice des
taux tissulaires,
3) à la capacité de l'organisme de maintenir
l'équilibre du Mg pour des administrations et des taux
tissulaires suboptimaux et
4) aux besoins accrus créés par la croissance,
le développement la réparation, le stress, les
déséquilibres alimentaires, et les facteurs
génétiques et de l'environnement.
Le deficit magnésique provoque plusieurs anomalies dans
la structure et le fonctionnement. Par contre, le Mg est
remarquablement inoffensif même lorsqu'il est
administré par voie parentérale, à condition
qu'il n'existe pas d'insuffisance rénale, et qu'il ne soit
pas administré rapidement par voie intraveineuse.
Ainsi, jusqu'à ce que des données
définitives soient disponibles pour les administrations
optimales dans différentes conditions physiologiques et
pathologiques, les administrations per os de vraient
être accrues jusqu'à 6-10 mg/kg/jour pour les
adultes jeunes, et à deux fois plus pour les sujets
subissant un anabolisme actif, ou pour ceux se trouvant sous
l'effet d'un stress.
Controversy and confusion still attend consideration of human
magnesium (Mg) requirements. Ten years after the First
International Symposium on Magnesium, it is necessary to repeat,
"The place of Mg deficit in medicine is still not universally
accepted" [269]. Magnesium is not unique in having its
requirements for maintenance of optimal growth, development, and
health poorly appreciated. Most to blame for physicians'
reluctance to accept the possibility that Mg deficiency might be
contributory to medical problems, is the classic concept that the
daily requirement of a nutrient is the amount that prevents overt
signs and symptoms of deficiency. In the case of Mg, plasma
levels are maintained within fairly narrow limits, despite losses
and needs that exceed supplies or absorptive and
retention-capacities. Hypomagnesemia, sufficient to cause
convulsions or dysrhythmias that are refractory to therapy other
than Mg, reflect depletion rather than deficiency. Yet even in
such instances, most clinicians attribute the efficacy of
Mg-therapy to its pharmacologic activity, rather than to repair
of a deficiency.
Lesser degrees of deficiency are recognized predominantly by
those who have demonstrated reduced erythrocyte Mg-levels and
Mg-responsiveness of patients with a variety of neuromuscular or
psychoneurotic syndromes [69, 70, 72, 74, 87, 88, 137, 138].
Metabolic balance studies have supported the contention that in
such patients, Mg-intakes from their self-selected diets are
inadequate to maintain Mg-balance [69, 73, 74]. With few
exceptions [87, 88, 138, 139, 277] except in Latin-language
countries, most physicians do not accept the likelihood of
marginal Mg-deficiency in neuromuscular or psychological
disturbances, in disorders of the alimentary tract or endocrine
system or as a possibly contributory factor to the early origins,
or early or later manifestations of cardiovascular, renal,
skeletal or immunologic diseases.
Marginally low plasma Mg levels, that might reflect more
significant tissue deficiency, are usually disregarded.
Difficulties in determining and interpreting cellular levels
limit attempts to obtain tissue Mg levels. Only in research
ventures are tissue Mg levels measured. Further confounding the
problem is the complexity of interactions that affect dietary
Mg-requirements. The metabolic balance technic provides the
baseline requirements data, usually as influenced by single
dietary variants under strictly controlled conditions. Invaluable
as a method of determining minimum requirements in a test
environment, this procedure is not readily adaptable to usual
living conditions, because of the multiplicity of factors that
affect requirements.
A 1964 analysis [275], of metabolic balance data collected
from studies done throughout the world, indicated that Mg-intakes
by adults, of less than 5 mg/kg/day, are unlikely to maintain Mg
equilibrium. Intakes of 7-10 mg/kg/day, particularly by young
men, are probably preferable [269]. After study of factors that
increase requirements, and consideration of the safety of orally
administered Mg to those without significant impairment of renal
clearance, the author reiterates that recommendation. Translating
this to total daily Mg intakes, 450-700 mg might be desirable for
a 70 kg man, and as much as 600-1000 mg/day for a 100 kg man
— whether much of his weight is fat or muscle. Young women
maintain equilibrium on lower mg/kg/day Mg-intakes [269].
Basing their recommendations largely on metabolic balance
studies, much lower intakes were recently officially recommended
in the United States [101]: 300 and 350 mg/day for young women
and men, respectively; 400 mg for 15-18 year old boys; 450 mg for
pregnant or lactating women. The low infant RDA (Recommended
Dietary Allowance) for Mg (70 mg/ day, or less than 10
mg/kg/day), based on the amount present in infant formulas [101,
307] urgently requires re-evaluation, in view of clinical
evidence of infantile hypomagnesemia — especially in
bottle-fed babies (Review: [271]), and early metabolic balance
studies that rapidly growing children sustain strongly positive
Mg-balance while on diets providing as much as 20 mg/kg/day or
more (infra vide). This evidence, that high Mg-intakes
result in retention of large amounts of Mg by subjects undergoing
growth and repair, suggests that the optimal Mg-intakes of
pregnant and lactating women, adolescents, athletes in training
and in competition, and convalescents, be fully studied.
That even the modest, officially recommended, Mg-intakes are
not met by most Americans and Canadians, has been shown by
dietary surveys [200, 292, 307, 324, 330]. Analysis of thousands
of typical sample meals of American children and adults of all
ages and both sexes, showed Mg-intakes to be below the RDA. Most
of randomly selected students in 50 colleges in the United States
[307] and one from Canada [292] chose meals that provided much
less than the RDAs for Mg and twice as much, or more, calcium
(Ca) and phosphorus (P) (Figure
1). This dietary pattern causes concern when considering the
straightline correlation of the increased incidence of ischemic
heart disease death rates in countries with high dietary Ca/Mg
ratios [158, 311], and the evidence that high P-intakes intensify
damage caused by Mg-deficiency [272, 272]. Other diseases,
notably gastrointestinal neoplasms and leukemias, have geographic
differences in distribution that match low Mg-availability [7,
270]. Animal and in vitro studies provide data that
suggest contributory roles of high Ca/Mg ratios [12, 13,
270].
Intervention studies are being implemented or planned in
Canada [336], and Finland, to determine whether Mg supplements,
given to high-risk individuals or population groups will reduce
the incidence of cardiovascular diseases (personal
communications: J. Marier, H. Karppanen). We should also
ascertain whether, and to what extent, Mg-supplementation of
infants would prevent Mg-responsive infantile irritability and
convulsions, to which high-risk infants and formula-fed infants
are more vulnerable than are breast-fed normal infants [267, 271,
274]. Whether such supplements can reduce the incidence and
severity of diseases with early and late manifestations
resembling those producible by experimental Mg deficiency, the
roots of which might be in early life [268, 271, 272], will
require much more investigation and evaluation. Whether Mg
supplements might protect against cardiovascular consequences of
diets too rich in protein, fat, and sugar, and in Ca, vitamin D,
Na and P will remain speculative until long-term intervention
studies can be extended and evaluated. In view of fiber's
reduction of retention of Mg and trace minerals, evaluation of Mg
requirements should be part of this approach to preventive
medicine.
Difficulties in establishing human magnesium needs
Factors that affect human nutrient requirements
There are individual, dietary, and environmental factors that
influence the amount of each essential nutrient that is needed to
maintain resistance to disease and optimal physical fitness [143,
337]. Under essentially normal conditions, in the developed
world, the host factors are the most important. Individual
differences: age, sex, genetic factors, activity, customs, and
responses to infection and other stresses, all affect nutritional
requirements. Dietary factors: chemical form and quantity of the
nutrient, levels of other nutrients, and food processing
influence the amount available to the consumer. Environmental
factors, such as water and soil, climate, season and altitude,
and stress factors, all influence either the availability or the
retention of the nutrient. Figure
2
Interactions among all of these factors influence nutritional
requirements of individuals, families, racial, or ethnic groups
and of these groups in different geographic areas. Epidemiologic
surveys, that correlate at least some of the nutritional variants
with differences in disease distribution patterns, provide clues
to the role nutrition can play in predisposing to, or protecting
against, pathologic processes. Intervention studies, in which we
change the intake of one or more nutrients that are suspected of
playing a role and evaluate subsequent changes in morbidity and
mortality of high risk-groups, provide further insight into the
efficacy of altering the nutrient intake.
Since Mg plays such an important role in protein and nucleic
acid synthesis [79, 253, 310, 318, 321, 322, 341] it is to be
expected that those undergoing rapid growth and repair have high
Mg-requirements. Primary Mg-malabsorption [222, 227, 228, 257,
294] and renal-wastage [104, 110, 209, 229, 237, 255], often
familial, indicate that there are genetic differences in
Mg-requirements. These abnormalities are probably at the ends of
bell-shaped curves of distribution of Mg-absorptive and
renal-retention capacities. The extent to which Mg requirements
are affected by genetic differences in utilizing nutrients (e.
g., Ca, Zn, or vitamins D, B1, B6, E, and
A) must be investigated. Of particular interest are
susceptibilities and dependence on nutrients — excesses or
deficiencies of which affect Mg-utilization. Whether one is
phlegmatic or excitable is at least partially genetically
determined, although environmental factors play a role. One's
physical and mental prowess, in conjunction with one's
competitiveness (to which genetics and environment contribute),
will influence the degree to which one seeks the stress of
excessive physical training and athletic competition, or
intellectual or business ventures that create psychological
strains, and can lead to overwork. Those whose work or preference
subjects them to crowding, and to noise, vibration and flickering
lights as in subways or discoteques — are also subjected to
stress and the release of the stress hormones: catecholamines and
corticosteroids. These hormones have long been known to mediate
both physiologic and pathologic responses to stress [17, 235,
236, 281, 283, 284]. Cardiac and renal damage is intensified by
Na, P, Ca, and calcemic agents. Potassium, Cl, and Mg-inadequacy
each worsens the lesions; the administration of each is
protective [17, 169, 171, 244-247, 281-284, 300, 301]. Less
widely appreciated is the increase in secretion or release of
corticoids that is caused by low Mg [41]. Catecholamine release
is also increased by low Mg and high Ca concentrations [30, 62,
236, 254]. Some of the pathways by which stress causes Mg loss,
and the conditioning to such loss by dietary imbalances, are
designated in Figure 3.
Metabolic balance estimation of nutrient requirements
A valuable investigative tool, metabolic balance
determinations require careful evaluation. Such studies provide
data on the amount of a nutrient that enters and leaves the body
during the collection period. They provide no information on
internal distribution or exchange. Directed towards establishing
normal (minimal) requirements under standardized conditions,
variables are limited [143]. Because the procedure is cumbersome
and expensive, and housing in the protected environment of
metabolic units is preferred, in order to obtain uncluttered data
on the influence of single (usually dietary) variants on the
utilization of the nutrient under study, the results are
generally based on responses of few subjects under artificial
stress-free conditions. Furthermore, young adults who volunteer
to live regimented lives for long periods, isolated from normal
outside activities, are likely to have relatively placid
temperaments. Metabolic studies done with volunteers living at
home or in dormitories, but who are willing to consume only the
monotonous diet provided in metabolic units, and to collect all
excreta, also are likely to comprise a selected group of
individuals who do not reflect a wide range of dispositions. Even
among such subjects, differences have been observed in the course
of long-term metabolic studies, without altering dietary or
environmental conditions [143].
Still another source of error is the pre-study nutritional
status. Those with severe inadequacies (which might cause
enzymatic dysfunction) might not adequately utilize high intakes
until the metabolic abnormalities are corrected. Thereafter,
positive balances should be maintained until the body deficits
are repaired. On the basis of evaluation of 40 years of metabolic
balance data from short-term and long-term studies, Hunscher
[143] pointed out that subjects can maintain equilibrium even
when their bodies are in actual debt, or below safe levels for
any age or stage. She asked, "should we be complacent in
accepting a lesser" than optimal "concentration in the body by
being satisfied with equilibrium of flow resulting in
unsaturation of tissues?"
Despite these defects, metabolic balance studies continue to
provide important data — particularly when used in
longitudinal evaluation of requirements during the course of an
illness or physiological process [24].
Metabolic balance studies of magnesium requirements
Adults
Many of the metabolic studies, from which the 6-10 mg/kg/day
magnesium intake recommendations for adults were derived [275],
were done with free-living volunteers rather than in metabolic
units. Such studies are subject to errors that result from
failures to adhere to prescribed diets and from losses of
samples. One may question whether the errors that result from
selection of those with personalities suitable for the voluntary
restrictions, and/or isolation from normal stresses, might not be
at least as great. The meticulous balance studies, from which
estimate of daily Mg needs of 300 mg or less have been derived,
probably indicate minimal requirements [24, 143]. As with other
nutrients, equilibrium can probably be established at sub-optimal
and optimal intakes [143] as body deficits are repaired. The
first clue that Mg supplements could produce strongly positive Mg
balance, even after equilibrium had been maintained at much lower
intakes, was provided by a long term study in a prison in 1926
[44] (Figure 4). One of two
men who had been in strong negative Mg balance for the preceding
two months, stored Mg during the weeks of supplementation
(intakes of about 12 mg/kg/day). The other, who had a history of
renal disease, merely lost less Mg. Perhaps he is the first
recorded case of renal Mg wastage, about half the amount of Mg
ingested daily, at low and high levels, was excreted in the
urine. The study of healthy young college men fed a high Mg
(8.9-11. mg/kg/day) liquid diet, such as was being investigated
for patients needing tube-feeding, showed strong positive
balances during almost all of the periods 44 week-long ( [113],
Fig. 5). One strong negative
balance occurred during a stressful week of final examinations. A
study of stable, ambulatory hospitalized men, who had been in
Mg-balance on a diet that delivered about 185 to 300 of Mg/day,
showed strongly positive balances early during Mg-supplementation
that increased Mg-intakes 4-fold [275]. The retention of Mg then
gradually diminished until equilibrium was again established
after several weeks of high Mg-intakes.
Other long-term studies confirm the inadequacy of diets that
provide under 5 mg/kg/day for men and suggest that even the RDA
for women might not be sufficient for equilibrium. Two men with
mean daily Mg-intakes (from self selected diets) of 4.2 and 1.8
mg/kg/day lost 40±35 and 90±40 S. E. mg Mg/day
[303]. Fifteen healthy young women, consuming a controlled diet
that provided 265-305 mg Mg/day (or 4-5 mg/kg/day), were in mean
strongly negative Mg balance over 3 consecutive 20-day-periods
(Figure 6) [148]. The diets
provided high Ca/Mg and P/Mg ratios, such as are customary in
American diets (supra vide). They had strongly positive
Ca-balances, which are not necessarily salutary, in view of soft
tissue calcinosis that occurs in experimental Mg-deficiency [82,
102, 128, 203]. Their negative P-balances, despite their high
P-intakes, are provocative, in view of the association of
hypomagnesemia with hypo-phosphatemia (Review: [161]. The authors
commented that the sustained Mg-losses on Mg-intakes considered
adequate, suggest that higher daily Mg-intakes of 385 mg (or
about 6 mg/kg/day) might be necessary to maintain a 140 lb (64
Kg) woman.
Adolescents and children
Relatively few Mg-balance studies are available for those with
rapid growth rates. Older adolescents (18 years) are commonly
considered with teen-age boys as having high nutritional
requirements. Girls that age are commonly considered with young
adults. Mg-requirements of growing, developing infants, children
and adolescents are greater than are those of adults — with
the exception of pregnant or lactating women, and of those under
stress or during convalescence.
Adolescent Girls (Figure 7): The first
metabolic balance study of adolescent girls, in which Mg was
measured, was a 1936 study [327] of 22 girls (11-15 years old),
eating self-selected diets that provided 6-10 mg/kg/d, or more
than the RDA for Mg. Eight of the 22 girls, studied for 6 days,
retained less than 10 mg of Mg/day — an amount that might
be insufficient to meet the growth and development needs of this
age group. On even lower Mg-intakes (190-195 mg/day, or 3.3-5.6
mg/kg/day), none retained more than 10mg/day of Mg. The only
girls not in negative balance were small, and thus received about
5 mg/kg/d [117]. In a subsequent study, in which Mg-supplements
were given to raise the average daily intake to 286 mg, there
were 9 balance periods with Mg-retentions of over 10 mg/day: 5
girls, when on a low zinc-intake, and 4 when on a high
zinc-intake [118]. It is noteworthy that the latter two studies
showed that girls on marginal magnesium-intakes retained less Mg,
when their zinc intakes were high. Low though the Mg-intakes
were, during these studies, they were actually higher than that
consumed by 15 14-16 year-old girls, whose 24-hour usual diets
were analyzed [330]. The mean Mg-intake was 128 mg, with total
intakes as low as 56 mg. Of 15 American girls, 17-18.8 years of
age, fed a controlled lacto-ovo-vegetarian diet that provided a
mean Mg-intake of 321 mg/day, only 7 retained more than 3% [198].
The investigators commented that this indicates that girls up to
18 years of age have the higher requirements of adolescents,
rather than the maintenance needs of adults. The higher than
customary fiber content of the largely vegetarian diet, however,
might have influenced the results, possibly decreasing
Mg-absorption (Infra vide).
Adolescent Boys (Figure 8) : Magnesium
requirements of adolescent boys have been correlated with their
protein intakes, because of their rapid rates of growth and
development [4, 263]. A study of boys who were 13-14 years old at
the outset, and who gained 6 to 13 kgs from the beginning to the
end of the study, which spanned two years, were in negative Mg
balance when both protein and Mg-intakes were sub-optimal [263].
Increasing the protein intake to 93 gm, an amount in excess of
requirements, improved the retention at the low intake of Mg.
However, it was cautioned that the Mg retained with the high
protein-intake, low Mg-intake, although probably adequate for
maintenance of soft tissue content, was unlikely to meet
Mg-requirements for growth and maturation. With the high
Mg-intake of 740 mg, with low protein-intake, there were strongly
positive Mg-balances the first year, but most had negative
Mg-balances the second year. This might indicate the greater
inadequacy of the 43 gm protein diet for the boys, the second
year, when they were bigger. Strongly positive Mg-balances were
maintained by most when both Mg and protein-intakes were high.
The authors concluded that, since there is experimental (animal)
evidence that a deficiency of Mg relative to protein is harmful,
whereas an excess of Mg relative to protein is probably not, Mg
requirements should be set to permit an adequate retention ratio
of Mg/N. They suggested that calculation of Mg requirements on
the basis of Mg/N retention equal to or higher than the Mg/N
ratio of the whole body, might yield a preferable figure for Mg
needs during growth, than is provided by balance studies alone.
Like the 17-18 year-old girls, whose balance studies indicated
that their Mg-needs were that of younger adolescents, rather than
the lower maintenance need of adults [198], with whom they are
usually categorized, 18-20 year-old men went into negative
balance on Mg-intakes of 3.5 -7 mg/kg/day [4]. At those low to
moderate Mg-intakes, changing the protein content of the diet
from low (48 gm) to high (141 gm) exerted no influence on
Mg-retention. When they were given 10.5 mg of Mg/kg/day,
equilibrium to strongly positive Mg balances were achieved in
half on the low protein diet, and in 5 of 6 when they were given
a high protein diet.
Infants and children
The amount of Mg needed by infants and children is not
certain. A review of the sparse metabolic balance data available
in 1940 [67], correlated with tissue Mg-levels and growth curves,
led to estimation of 10-20 mg/kg/day as the Mg-requirement. The
optimal Mg-retention during growth and development has not been
elucidated.
Infants: Infant Mg requirements
depend upon the adequacy of maternal, and thus of fetal supplies
whether the infants are premature, small for gestational age
(SGA), born after complicated gestation or delivery, or whether
they are full-term and healthy. For baseline data, we must
ascertain how normal infants respond to normal milk diets, how
supplements influence their Mg-retention, and how the mineral
retention of breast-fed infants differs from that of those fed
cow's milk formulas or formulas adapted to resemble human milk.
During the first week of life, normal, full-term infants, whose
Mg-intakes provided 3.5-4.4 mg/kg/d from human milk, and 6.8 to
10 mg/kg/d from cow's milk formulas adapted to resemble human
milk, retained small amounts of Mg [189, 287, 331 332]. Almost
5-fold more Mg was retained from cow's milk. However, infants at
all ages retained far more Ca and P when fed cow's milk, as
compared with those fed human milk, a phenomenon termed
"supermineralization" of cow's milk-fed babies [299]. Despite the
retention of large amounts of minerals by cow's milk-fed babies,
they are much more subject to hypomagnesemic hypocalcemia in
association with hyperphosphatemia than are breast-fed babies
[45, 271, 272, 206]. The much higher mineral content of cow's
milk undoubtedly is the major factor in the high mineral
retentions of formula-fed infants. The addition of vitamin D is
also contributory. Metabolic balance studies have shown that
vitamin D-supplements have little or no influence on Mg-retention
of human milk-fed infants, but profoundly increase Ca- and
P-retention of cow's milk-fed babies [285, 299, 331]. One of the
studies [285] showed that infants given no vitamin D or up to 200
I. U. were in positive Mg balance during 44 metabolic study
periods, and in negative balance during 8 periods. In contrast
the Mg balances, of those who were given more than 200 units of
vitamin D daily, were positive during only 16 of the test periods
and were negative in 9. Breast-fed neonates showed slight rises
in serum Mg in the first week, whereas vitamin D-free formula-fed
infants showed a slight fall [111]. Adding 600 units of Vitamin D
lowered the serum Mg further in bottle-fed babies, but not in
infants on mother's milk. Administration of the vitamin D
metabolite, 1-25-(OH)2D3 raises the total
and ionized Ca low-birth-weight infants during their first 48
hours, without affecting plasma Mg [42]. Such infants, who are
likely to have low Mg-stores, are commonly treated with calcemic
agents, which increase Mg-requirements [271], not necessarily
reflected by low plasma Mg levels.
An early long-term study of infants fed formulas, without and
with added fiber (celluflour) or spinach purée (which
doubled their Mg-intake) showed some interference, by fiber, with
Mg-retention [262] (Figure 9).
When the amount of Mg in the milk formula was doubled, by adding
mineral salts equivalent to those in the spinach, all four of the
infants retained more Mg. This study is noteworthy for two
reasons: it shows that on Mg-intakes of 50-75 mg/day from cow's
milk formula (the current RDA for young infants), the infants
retained only enough Mg to remain in equilibrium — a
scarcely desirable situation for rapidly growing babies.
Furthermore, it shows that they did not retain the extra Mg
provided by the spinach when they were under six months of age.
This provides further justification for the recent observation
that early infant-feeding of solid foods (as has been the
practice for the past half century) is not desirable [168].
Children: Early metabolic balance
studies of Mg retention by 5 to 9 year-old children, consuming
diets providing about the RDA of 8-10 mg/kg/day to as much as 25
mg/kg/day, showed that negative balances to retentions of no more
than 10 mg/day were the general rule [56, 57]. One study showed
that children on 10mg/kg/day or less were in negative balance;
most of those receiving over 18, to as much as 40 mg/day of Mg,
retained very large amounts of Mg (120 to 440 mg/day) during
their 4 to 5 day balance studies [231]. Whether such retentions
can be verified, and how long they would persist, would provide
important information as to optimal Mg nutriture of children.
Subsequent studies have shown that young children given diets low
in Mg (4-9 mg/kg/d) maintain Mg-equilibrium or retain about
20mg/day [125]. A study in which the same children were given
diets providing different levels of Mg, showed strong
Mg-retentions on the high (13-18 mg/kg/day) versus the low (<
10 mg/kg/ day) intakes.
Anabolic processes requiring more magnesium
Those forming new tissues, whether as a result of growth and
development, athletic training, convalescence, or pregnancy and
lactation, have increased nutritional needs. The supply of
magnesium, which plays important roles in nucleic acid and
protein synthesis and in numerous enzyme-functions [128, 253,
310, 318, 321, 322], often insufficient to maintain equilibrium
during stable phases, can fall to critical levels during growth,
development, and repair processes. Further study is needed to
define how much Mg is necessary for optimal function at
those times.
Pregnancy and lactation
The condition, during adult life, in which Mg-requirements
have recently recognized as being elevated (RDA = 450mg/day) are
pregnancy and lactation [101]. Nevertheless, pregnant women whose
Mg-metabolism has been studied, selected diets that provide less
than half that amount [14, 144]. Negative Mg-balances have been
found in middle-class pregnant American women [14]. Review of the
literature shows that the Mg-intake during pregnancy has been
declining since the turn of the century [271]. The first report
found was of a 1914 study in Germany [164] showing that on daily
Mg-intakes of 338-510 mg, pregnant women retained from 97-159
mg/day in the last trimester. Longterm studies of pregnant women
in Finland [136] and in the United States [149, 141] showed that
in Finland, where the Mg-intake during pregnancy was 177-389
mg/day, there were many negative balance periods (Figure 10). A healthy American
quadripara, whose Mg-intake during the last half of pregnancy was
high (590-615 mg/day), remained in strongly positive Mg-balance
during her last trimester [141]. An American teen-aged primipara
with a poor nutritional history and whose Mg-intake was below the
RDA, retained less Mg during the last two lunar months of her
pregnancy [140]. It is worth noting, here, that adolescent
mothers, whose own Mg-needs are likely not to be met, are
particularly at risk of gestational Mg-deficiency.
The optimal Mg-intake during gestation remains to be
determined, but there are several findings that suggest that
gestational Mg-inadequacy might be more common than realized.
Pregnant women tend to develop somewhat lower serum Mg-levels
than are seen in age-matched non-pregnant women [61, 121, 221].
Correction for hemodilution has shown the hypomagnesemia to be
real during the first half of pregnancy and in the last month
[61]. Hypomagnesemia has long been recognized in eclampsia [1,
121, 126]. Mg-therapy has been used to control pre-eclampsia and
eclampsia since the first quarter of this century [167]. On
occasion, investigators have wondered whether the similarity of
some of the toxemic manifestations to those of Mg-deficiency
might indicate that the improvement on Mg-therapy might reflect
repletion of a deficit [75, 98, 121, 263, 271, 274]. It is also
possible that Mg-inadequacy might also contribute to gestational
hyperparathyroidism, which is so common as to be termed
"physiologic" [54]. The occurrence of hypomagnesemia,
hypocalcemia, and hyperparathyroidism, especially in the third
trimester [28, 54, 61, 121, 221, 329], suggests that
hyperparathyroidism might result from the mineral inadequacies,
rather than be physiologic [267, 271]. In fact, the development
of infantile hypocalcemia is considered an indication of need to
test the mother for hyperparathyroidism [ 18, 105, 124]. Maternal
hyperparathyroidism has been detected in mothers of infants with
neonatal hypomagnesemic hypocalcemia [60, 85, 213].
The vulnerability, during the third trimester, to gestational
abnormalities to which magnesium inadequacy might be
contributory, might reflect the fetal needs for Mg during that
time [48, 271, 333, 334]. It has been estimated that the daily Mg
content of the fetus doubles during the ninth and tenth lunar
months [48, 333]. Perhaps the improved perinatal salvage rate
(10% versus 25-35% fetal loss) of infants of eclamptic women
treated with Mg, as compared with those otherwise treated [234,
343, 345] might reflect meeting of fetal needs.
Recovery after starvation
Sudden cardiac deaths have been reported during total
starvation [52, 107, 291], during refeeding after total
starvation [107] and while dieting on a liquid protein
preparation [207]. The latter patient had a low serum magnesium
level (1.5 mEq/L) that did not rise to more than 1.67 mEq/L
despite an infusion with Mg, and she died of ventricular
fibrillation and cardiomyopathy. Because short- and long-term
fasting of non-obese volunteers [47, 298] and obese patients [64,
65] results in substantial Mg-losses that are not always
associated with low serum Mg levels, attention should be paid to
meeting Mg-needs both during fasting and re-feeding.
It has been shown that malnourishment, with resultant
metabolic abnormalities, interferes with adequate utilization of
nutrients which is reflected by delays before positive balance
(e. g., of Ca) is achieved with repletion [243]. Evidence that
this holds true for Mg was provided by the study of Mg-balances
of starved patients, early after admission to the hospital and
during Mg-repletion [205, 206] (Figure 11). Strongly negative Mg
balances persisted on diets providing 800 mg Mg/day. Supplements
(Mg lactate), that increased the daily Mg-intake to 1-1.36 g/day
produced retention, but losses recurred when the Mg-rich diet,
alone, was fed. The investigators reported that subnormal
Mg-retention was only gradually corrected during the half to one
year period of observation. Possibly such patients are in need of
very high Mg-intakes. Noteworthy are the essentially normal serum
Mg levels in 21 starved patients, which fell to sub-normal levels
during the first 12 weeks of re-feeding and Mg-repletion, and
rose to normal in most only after a half-year of
convalescence.
The Mg-deficit of starvation is contributed to by the
associated increased production of lactic acid and lactate, and
by ketone acidosis — conditions that increase urinary
Mg-loss [19, 153, 180, 181]. In such instances, Mg-deficiency can
be masked by the egress of tissue Mg, whether the metabolic
abnormality is caused by decompensated diabetes mellitus [201,
202, 218], starvation [64, 65, 298], or the malnutrition of
chronic alcoholism [156]. The stress of starvation or of
alcohol-withdrawal is another factor in Mg-depletion. The stress
hormones cause Mg-loss directly and as a result of lipolysis,
with release of free fatty acids [239, 241] (Figure 3). During the acute phase of
alcohol-withdrawal, free fatty acids increase in the plasma
[204], with resultant decrease of serum Mg [93, 95, 96]. Starved
obese patients have ample fat stores to yield free fatty acids;
those taking liquid protein diets to lose weight have an added
risk factor: the increased urinary Mg-excretion produced by
protein loads [68, 180].
When patients are malnourished as a result of starvation or
disease, and are refed without consideration of the high Mg-needs
to make up for tissue-losses and meet the requirements for new
tissue-formation, there can be serious consequences [339].
Children with protein-calorie malnutrition, for example,
deteriorated rapidly and developed cardiac complications when Mg
was not provided during the high caloric-protein treatment [33,
34]. Similarly, the malnutrition and vitamin (B1)
deficiency of alcoholics can be refractory to therapy until the
Mg losses are repaired [339].
Magnesium requirements of the aged
Metabolic balance data are lacking for aged people, but there
are fragmentary findings that suggest that their Mg-requirements
are likely not to be met by their diets. A survey in Belfast
shows that the diets of institutionalized and
non-institutionalized old men and women provided from 160 to 230
mg/day [312]. The decreased intestinal Mg-absorption by old, as
compared with young normal subjects and patients [154, 216] puts
them into further Mg-deficit. Serum Mg levels of aged subjects
have been reported to be lower than in young adults [131, 245]
and to be about the same [159, 304]. Since the Mg-retention of
aged mice is much less than that of young mice [63], and the life
expectancy of rats kept on low Mg-intakes for their entire lives
is reduced [133], definition of Mg needs of aging people, and the
effects of Mg-supplementation seems worth exploring.
Dietary factors that increase magnesium requirements
Many nutrients increase Mg-requirements when supplied in
excess (Reviews: [71, 72, 76, 166, 180, 269]). These include
calcium and calcemic agents like vitamin D, phosphate, phytate,
sodium potassium, protein, carbohydrate and fat. Emphasis on the
role of saturated and unsaturated fat on cardiovascular disease
[8, 9] has led to recommendations for major dietary changes that
have been questioned [100, 184, 296]. Generally disregarded is
the effect on Mg-utilization of the dietary fat. The newer
nutritional approach to hyperlipidemia and atherosclerosis
— that of increasing the fiber content of the diet [171,
217, 308] is one that interferes with Mg-absorption (infra
vide). Since the availability of Mg has clearly been shown
to affect the degree of cardiovascular damage in many
experimental models (Review: [244, 271, 273, 278, 279, 300, 301])
its availability in diets that alter fat and fiber intakes might
well be critical in influencing the prophylactic efficacy of such
diets.
Effects of dietary fat on magnesium
requirements
As early as 1918, it was shown that fat interfered with
Mg-absorption [261]. Mg was better absorbed from meals that
provided 436 mg/day than it was from meals that provided 314 mg
Mg/day and that were rich in butter fat. The two children (5 and
8 years of age) were in strongly negative Mg balance having
3-days of high-fat diet. In another early, short-term balance
study [29], substitution of butter for margarine, in a diet rich
in Mg (800-832 mg/day), resulted in slightly increased
Mg-retention by each of 4 young women. In two, negative balances
were converted to positive. Linoleic acid has also decreased
Mg-utilization by young men [125, 149] and women [148] fed diets
much lower in Mg. Mg-retention fell as the linoleic acid content
was increased. At Mg- intakes of 4-6.3 mg/kg/day, or 300-370
mg/day, most of the young men were either in Mg-equilibrium or
slightly negative balance on controlled diets providing 9-10% of
the free fatty acid; Mg-losses increased as the linoleic acid
content was increased to 30% of the calories. Young women, who
lost an average of 63 mg Mg/day while on a controlled diet that
provided 4.2-5.4 mg Mg/kg/day (supra vide, Figure 6), showed rising blood lipids,
even though the dietary fat was low (1 g/day) (Figure 12). This observation is
analagous, though lesser in degree, to the experiments that
showed that Mg-deficiency intensified the hyperlipemia of animals
fed hyperlipemic, atherogenic, thrombogenic, or glucose-rich
diets [129, 142, 249, 259, 260, 280, 313-315]. Of particular
interest is the increase in levels of β- and pre-β
lipoproteins in Mg deficient rats, and in pigs [224], that fell
with Mg-supplementation, since preliminary clinical studies
suggest a similar effect of Mg-supplements in patients with
hyperlipemia [127, 178, 226, 293]. Since increasing the
Mg-content of cardiovasopathic diets has protected animals
against tissue lesions [224, 259, 260, 271, 279, 289, 300, 301]
members of high-risk families and populations, who might have
high Mg-requirements, might benefit from Mg-supplementation.
Diets designed to meet official recommendations to lower the
total intake of fat to 35% or less of total calories in order to
reduce the risk of cardiovascular disease, provide less than the
RDA for Mg [200]. Possibly the prophylactic effect of such diets
might be improved by correcting sub-optimal Mg-intakes. The
optimal Mg-intake of those with abnormal handling of fat and who
are vulnerable to cardiac and arterial disease requires
study.
Effect of fiber and phytates on magnesium
requirements
The incidence of many chronic diseases is lower among
population groups consuming diets rich in fiber than among those
eating refined foods low in fiber. This observation led to the
official recommendation [308] that Americans increase their
intakes of fiber. Careful scrutiny of findings from early
metabolic balance studies discloses much valuable information
that complements the newer work which shows that fiber causes
losses of minerals, including Mg. The effect of phytate on Ca, P
Mg and trace minerals has attracted interest. The first long-term
metabolic study, that compared the influence of bread made of
white or brown flour, with and without phytate or Mg (added to
the white flour) showed that phytate interfered with the
absorption of the 2-fold greater amount of Mg in the brown bread
[186, 187]. When the same amount of Mg (as carbonate) was added
to white bread, its retention was improved. A subsequent study
[323], in which balances were determined in two men, studied for
14-15 consecutive weeks, on diets with low- and high-fiber
breads. They showed initial profound Mg-losses when a pound of
high-fiber bread was substituted for other food (Figure 13). The retention of Mg
improved over the 7 and 8 weeks on that diet. They were in
Mg-equilibrium when white bread was substituted, despite the
smaller amount of Mg eaten. All of the 12 Ceylon medical students
on a controlled diet resembling what they usually ate: largely
brown and white rice, vegetables, and moderate amounts of animal
protein [53] were in Mg-balance (Figure 14). When white rice was
replaced by brown rice, they all retained less Mg, five going
into negative balance over the 5-week study. Providing only white
rice resulted in stronger positive Mg-balances, especially during
the weeks of consumption of a variety of rice that was rich in
Mg. Women consuming uniform diets low in Mg and fiber, or with an
adequate Mg intake provided by oatmeal, showed differences
depending on their weight [125]. The small women tended to remain
in equilibrium, or lost less Mg on the low-Mg diet or on the
phytate-rich diets, than did the larger women. The marked change
in balance in the same period is notable, particularly among the
women whose Mg-intakes were below 4 mg/kg/day on the low-fiber
diet, and whose intakes were not high on the high-fiber diet.
Mineral balance studies, undertaken more recently because of
zinc deficiency in the Middle East, that is related to high
fiber-interference with its absorption [232]. have shown that
whole grain breads also interfere with Mg-absorption [40, 243].
During 2 20-day metabolic balance periods, 2 young Iranian men
were in Mg equilibrium or slightly negative balance when half of
their food energy was derived from flat bread made from refined
flour [243]. When their bread was made with unrefined flour, each
went into negative Mg balance (44 and 129 respectively) despite
the two-fold greater Mg content of the whole grain bread. A
comparable longer-term study of 2 young Americans showed that
initially strongly negative Mg balances became positive or less
strongly negative when flat bread made from refined flour was
substituted for whole grain flat bread [40]. No adaptation to the
high fiber diet was observed in the study in which consecutive
balance results were reported [243].
Fiber added to controlled diets has also interfered with
Mg-retention. Studies of adolescent and pre-adolescent boys who
were given diets containing 420 or 336-333 mg Mg/day, showed that
both hemicellulose and cellulose increased the negative Mg
balance that was observed during most control periods [66, 194].
Similarly, men [160] and young women [288] retained less Mg when
fiber was added to their diets or when they consumed natural
fiber-rich diets. There was not a consistent trend toward
decreasing losses in six consecutive 5-day periods in the natural
fiber-rich diet. Noteworthy, is the lack of correlation of plasma
Mg levels with retention of Mg [288] (Figure 15). Most of the studies with
diets of added fiber gave less protein than is consumed by most
Americans. A more modest intake of fiber, added to a diet richer
in protein, has exerted little effect on magnesium retention by
men living in a metabolic unit and consuming uniform diets
providing about 300 mg/day [258].
Other nutrients that increase magnesium requirements
Since Mg is important in carbohydrate metabolism and in
protein and nucleic acid synthesis, those consuming diets rich in
proteins and sugars have high Mg-needs. Contributory to the risk
of Mg-inadequacy of such privileged people is the increased
urinary excretion of Mg that follows protein and glucose or
lactose-loads (infra-vide). Those whose high
protein-calorie diets are supplemented with large doses of
vitamins and minerals are at further risk of Mg-deficiency.
Protein: The physiological state and
activity, and the prior nutritional status, influence the effect
of protein-intake on Mg-needs. During protein-synthesis and
formation of new tissue by growing and developing children, by
athletes-in-training, by pregnant or lactating women, and by
those recovering from starvation or wasting illness, high-protein
diets increase Mg-needs.
Young adults, consuming controlled diets that provided 438 to
605 mg Mg (6.5 to 8.6 mg/kg/d) and moderate (45-70 g) or high
(100-130 g) of protein, absorbed and retained less Mg on
low-protein than on high-protein diets [188]. Of the 3 young men,
whose Mg-intakes were 7-8 mg/kg/d, 2 were in negative balance
(when sweat loss was considered) on the low, but not on the
high-protein diet. The young woman in that study, whose Mg-intake
was 5.8-6.5 mg/kg/day, was in positive balance on both protein
diets. In another study [294] of the effect of protein on Mg
absorption of young women, whose controlled diets provided more
Mg (5-7 mg/kg) than the RDA, all 10 were in strongly positive Mg
balances on low protein intakes (47 g protein). Providing 30 g of
animal protein to a basal controlled low-protein diet, fed to
young men (which increased their Mg-intake from 303 to 362
mg/day), or adding 100 mg of Mg as MgCO3 to the basal
diet, converted the negative Mg balance in 5 of 6 men to positive
[195]. Young women on low Mg intakes (2.5-4 mg/kg/day) were in
strongly negative Mg-balance on the very low protein-intake of 20
g, in Mg-equilibrium when the dietary protein was increased to
30-34 g, and in positive balance (+ 24) on a protein intake of 48
g [145]. Young adolescent boys retained more Mg from a low
Mg-diet (240 mg or 3.4-5.4 mg/kg/day) when the protein intake was
increased from 43-93 g (263, Figure 8). When the Mg-intake was
adequate for growth and development (740 mg, or 9.6-16 mg/
kg/day), the protein intake exerted less effect on the Mg
balance. Older teen-agers (18 20 years of age) tended to remain
in negative Mg balance on low and high protein intakes, when
their Mg-intakes were below 10 mg/kg/day, but most went into
positive Mg-balance on diets rich in both Mg and protein (4,
(Figure 8)).
In brief, the studies indicate that when intakes of both
protein and Mg are low the Mg balances tend to be negative.
Increasing dietary proteins to levels of "low protein" diets,
from extremely low levels, improved the Mg-retention of young
women, whose Mg-intake was kept low [145]. However, 18-20
year-olds on low Mg-intakes lost more Mg when their dietary
protein was increased from low to high [4]. Young adolescent boys
receiving sub-optimal amounts of Mg retained more Mg when their
protein-intake was increased from low to normal [263]. At
marginal Mg intakes (equal to the RDA), young men also retained
Mg better on normal than on low protein diets [195]. On high
Mg-intakes most of the balance periods of men and women [188],
and of young and older adolescent boys [4, 263] were positive,
whether the protein intakes were low or high. Additional to the
improved utilization by boys on the high Mg, low or high
protein-diets, as compared with that on the low Mg-diet [263],
the retention of Ca was improved [264]. Nitrogen balance was
about the same on the low-protein diet, on the high and low
Mg-intakes, and somewhat better on the high-protein diet when the
Mg-intake was also high [263]. The protein intake generally being
high, and the Mg-intake low in countries with diets resembling
that in America, it would seem advisable to increase the
Mg-content of the diet, or to give supplements so as to provide
6-10 mg kg/day to adults and twice as much to those with active
anabolic processes or under stress.
On the other hand, protein-loading has acutely increased
urinary Mg excretion [68, 180, 181]. Administration of 50 g of
gelatin (additional to the subjects' customary diet) resulted in
moderately increased urinary Mg-output of 12 to 24 mg/day over
the 6-day study [68]. Ingestion of 50 g of casein by 4 men
increased their mean renal excretion rates of Mg from 3.3 to 5.3
Eq/minute [181]. This effect of protein loads might have been
contributory to the hypomagnesemia of a patient who developed
fatal arrhythmia during feeding after a liquid protein diet for
obesity [210]. The recovery syndrome of children with
protein-calorie-malnutrition, during protein-calorie refeeding,
includes cardiac and nervous system disturbances, that can be
prevented or treated by addition of Mg to the diet [33, 34]. In
view of the direct and indirect evidence that high protein
intakes increase Mg-needs, its status should be evaluated in
those synthesizing protein and building new tissues, particularly
those at risk of prior Mg-inadequacy or loss. When such
evaluation is not feasible, providing such patients with at least
as much Mg as is required by young adolescent boys (>10 to 16
mg/kg/day [263] would seem to be the prudent course.
Effects of sugar, alcohol, and salt on magnesium
requirements: Diets in America and in other
industrialized countries are generally high in sugar and salt;
moderate to heavy alcohol-consumption is common. Thus, the
studies showing that each causes Mg-loss is germane to
calculations of Mg-needs.
Glucose: An oral glucose dose of 100
g caused almost as much urinary Mg-excretion by healthy young
men, as did an ample breakfast of cereal, milk, egg, ham, bread,
butter, jam and tea, or as did an amount of Mg and Ca equal to
that in the meal [135]. Glucose interferes with renal tubular
reabsorption of Mg [173-176, 180]. Lactose and galactose also
increase magnesiuresis [19, 180]. Oral glucose-tolerance tests of
normal children and adolescents (1.75 g glucose/kg to a maximum
of 75 g) caused a mean 9% decline of serum Mg on the first half
hour, that remained 5% below control values by the 4th hour
[249]. This effect was attributed to glucose-enhancement of
cellular uptake of Mg in normal children, that was not as
efficient in those with pre-clinical or clinical diabetes. These
clinical studies of the influence of acute glucose loads on
Mg-excretion in urine, and the laboratory animal evidence that
glucose (and insulin) increase tissue Mg-uptake [2], support the
observation that diets rich in sugar increase Mg-needs [72, 76,
166].
Alcohol: The severe Mg-depletion
caused by chronic alcoholism is well recognized, although not
always treated [92, 94, 97, 156]. Even moderate amounts, taken
before and with meals, increase urinary Mg-loss [157, 190, 193],
thereby increasing Mg-needs.
Sodium: Glucose loads augment the
renal tubular reabsorption of Na, at the same time as they
decrease Mg- reabsorption [175, 176]. Greater Na-loads such as
are provided when saline is used to expand the extracellular
volume, decrease serum Mg levels and increase urinary Mg
excretion [26, 134, 211, 325]. Fasting obese patients, whose Mg
losses exceeded the amount calculated to have been derived from
catabolized lean tissues, had only slightly lower than control
serum Mg levels, unless they received Na Cl supplements [64].
Those given 45 mEq Na daily exhibited as much as 25% drops in
serum Mg by the 20th-30th day of fasting. The reciprocal of this
finding is the Na and water retention seen in Mg-deficiency, and
the loss of Na (and water) produced by Mg-repletion [136, 120,
230].
Calcium and Vitamin D: Ca-excess,
such as has intensified lesions of Mg-deficiency in experimental
animals (Review: [271] is not characteristic of the human diet.
In the United States and Finland, where consumption of milk and
cheese is high, where (in the U. S.) milk is fortified with
therapeutic amounts of vitamin D, and taking vitamin-supplements
is common, the calcemic effect of vitamin D, which favors Ca
intestinal absorption and renal tubula reabsorption over that of
Mg [325], might make critical the Mg-inadequacy of the usual
American and Finnish diet [158, 268, 271, 311]. The
hyperlipidemia caused by even moderate vitamin D excess, or by
hyperreactivity to vitamin D [86, 182, 183, 271] might further
increase Mg-requirements.
Phosphates: Excess
PO4-intake is common in the industrialized world,
particularly among those consuming large quantities of colas
[185]. It is not uncommon for the PO4-intake to exceed
the RDA 2 to 3-fold (Figure
1). Such high intakes of PO4, particularly among
those eating highly salted foods and whose Mg-intakes are
marginal or low might well increase the risk of heart and bone
damage (Review: [271]). Are such dietary imbalances the human
analogue of the experimental cardiac necrosis produced by
Na2HPO4 which is protected against by Mg
and KCl (Reviews: [16, 171, 273, 279, 283])? When high vitamin D,
fat, protein and sugar content of the diet is added, a
cardiovasopathic diet that causes spontaneous myocardial
infarction in several animal species [244, 289, 300, 301], and
against which increasing the Mg content of the diet 5-fold has
been protective [247]. High dietary PO4 interferes
with Mg-absorption (Reviews: [128, 166]) and increases the amount
of Mg needed for normal function and survival by experimental
animals [32, 102, 214, 215, 225]. Thus the amount of Mg required
by those consuming diets rich in PO4 deserves
investigation.
Other vitamins, zinc, and magnesium needs
Abnormalities in metabolism of vitamins B6 and
B1 whether caused by genetic disorders or by diseases
that cause malabsorption, or whether caused by malnutrition, can
result in abnormalities of enzyme systems that are dependent,
also, on Mg. Vitamin E and zinc are additional nutrients with
interrelationships with Mg that might influence
Mg-requirements.
In these days of use of high doses of vitamins and trace
minerals, it is important to determine the effect of such
"megadosage" of trace nutrients on the macromineral — Mg
— that is usually consumed in suboptimal amounts. Overuse
of water-soluble vitamins is generally less of a problem than is
overdosage with fat-soluble vitamins, which are stored instead of
being excreted in the urine. However, excesses of even readily
eliminated nutrients can create difficulties if imbalances
result.
Vitamin B6:
Interrelationships of Mg and B6 deficiencies have been
recognized since it was found that the acute Mg-deficiency
syndrome was more rapidly produced in rats that were also
deficient in vitamins B6 (and B2) than in
those deficient only in Mg [114, 115]. The similarity in the two
deficiency disorders was also noted early [172]. Experimental
B6 deficiency has caused loss of tissue Mg [3]. It has
been associated with temporary hypermagnesemia and then
hypomagnesemia as the tissue Mg is depleted [15, 70, 248].
Several of the enzymes that require pyridoxal phosphate as a
coenzyme, such as pyridoxal phosphokinase, kynureninase,
transaminases, amino acid decarboxylases, transaminases, and
cystathionase, as well as Schiff base formation with amino acids,
also require Mg [15, 172, 191, 310, 318].
It is, thus, not surprising that Mg deficiency and
B6 deficiency produce comparable clinical disorders,
or that treatment with either Mg or B6 or both, have
been found effective in several clinical conditions in which one
or the other of the nutrients has been investigated. For example,
infantile convulsions have been caused by an infant formula
deficient in B6 [25], and by hypomagnesemia (Review:
[271]). There is evidence that a genetic vitamin
B6-dependent disorder that is characterized by
convulsions, is caused by interference with the glutamic acid
gamma-aminobutyric acid (GABA) system [266, 305]. The enzymes
involved are amino acid decarboxylases and transaminases, that
require both B6 and Mg for activation. B6
therapy is specific; the effect of Mg has not been studied in
this disorder. Three additional B6-dependent diseases
with central nervous system damage (mental retardation) are
cystathionuria, homocystinuria and xanthurenic aciduria [106].
The B6 enzymes involved are cystathionase and
kynureninase both of which also require Mg [172]. Since the
response of these diseases to B6-supplementation is
incomplete [106], it may be that Mg-requirements are also high.
B6-dependent anemia [106] might also be Mg-dependent.
The role of Mg deficiency in erythrocyte survival having been
clearly demonstrated [46, 81, 82, 84].
Whether pyridoxine and/or Mg-deficiency during pregnancy can
contribute to infantile B6-dependent diseases has not
been determined. However, B6-depletion during
pregnancy has been reported for many years [31, 316, 317].
Although Mg is a favored treatment for eclampsia, the proposed
role of Mg deficiency in the pathogenesis of gestational and
fetal abnormalities [267, 271, 274] has not gained as wide
acceptance. The B6-dependent disorders having a high
familial incidence, it should be possible to test the hypothesis
that meeting unusually high requirements of both nutrients might
correct the chemical abnormalities, and might be more effective
in the clinically manifest disorders than correcting only one of
the deficiencies.
The clinical efficacy of a combination of pyridoxine and Mg
[109, 233], and of Mg alone [154, 163, 207, 208], has been seen
in patients with calcium oxalate urolithiasis. It seems plausible
that the metabolic abnormality that results in such urinary tract
stones in geographic areas with low Mg-content of drinking water
[163], might be one in which there are higher than RDA Mg-needs.
The enhancement of clinical improvement by the use of both
B6 and Mg [109, 233] supports the premise of conjoint
activity. However, there is experimental evidence that high
B6 can intensify Mg-deficiency [162].
This finding points to the need for further study, and to the
possibility that megavitamin therapy might increase
Mg-requirements.
Vitamin B1: Mg-deficiency
interferes with response to thiamine in B1-deficient
rats [150-152, 338, 340] and in alcoholic patients, a not
surprising finding in view of the Mg-dependence of enzymes
requiring B1 (Review: [310] A genetic metabolic
disorder of B1 metabolism, subacute necrotic
encephalopathy (SNE) is similar to Wernicke's
encephalopathy, and might respond better to Mg plus B1
than to B1 alone. An infant with this disorder has
responded somewhat better to B1 when Mg was added to
the regimen (J. Cooper, personal communication). Another
infant has been found to have impaired B6 metabolism
[78], a further clue to a possible interrelationship with an
abnormality in Mg metabolism.
Administration of thiamine to B1 + Mg deficient
animals has intensified Mg-deficiency [150, 339, 340] and has
increased the blood levels of serotonum [151, 152]. Long-term
high dosage B1 therapy is common in chronic
alcoholics, who have Mg-deficiency and high Mg-needs, and in
those self-medicating themselves with megavitamins, whose
Mg-needs might thereby be increased.
Vitamin B2: There are
only few data on Mg/B2 interrelationships. Diets
deficient in B2, B6 and Mg or in
B2 and Mg caused more signs of Mg deficiency on higher
Mg-intakes than when only the Mg was deficient [114, 115]. On the
other hand, a high intake of riboflavin increased the
susceptibility to electrical stimulation of Mg-deficient rats
[212], thus high dosage B2 supplementation, whether
during recovery from alcoholism or by vitamin-faddists, might
increase the risk of Mg-deficiency.
Vitamin E: Experimental vitamin E
deficiency has lowered tissue levels of Mg in calves [27], rats
[90], and rabbits [342], and has precipitated lesions of Mg
deficiency in rats [112]. Administration of Mg has prevented
signs of E-deficiency in rats [265]. No data have been found on
the influence of high dosage vitamin E on Mg-requirements in
man.
Zinc, Vitamin B6 and Mg:
Both Zn and Mg are required for pyridoxal kinase [191].
Pyridoxine is necessary for tissue Mg-uptake [3] and might also
be for that of Zn, though there are different laboratory findings
[108, 139]. High oral doses [295], but not intravenous doses
[180] have decreased Mg-retention. Metabolic balance studies of
adolescent girls showed less Mg-retention from marginal Mg-diets,
when the dietary Zn was high than when it was low [117-119]. High
Zn-intake did not adversely affect Mg-balance when Mg-supplements
raised the daily intake to over 10 mg/kg. More attention is now
being paid to Zn deficiency, even in developed countries [23,
122], and Zn-supplementation, prescribed and taken without
medical supervision. The influence, on Mg-needs, of high
Zn-intake, should thus be ascertained.
Discussion and conclusions
There is a substantial gap between the typical Mg-intakes of
young Americans and even the RDA, which is based on studies
designed to determine minimum requirements. When the many factors
that increase Mg-requirements are considered, one can conclude
that absolute or conditioned Mg-inadequacy is common in the
industrialized world. Epidemiologic evidence that soft water, and
diets with high Ca/Mg ratios increase vulnerability to
cardiovascular disease [10, 11, 51, 99, 197, 219, 220] indicates
that when the Mg-intake is low, that provided in drinking water
can be critical. Nonetheless, determination of the Mg-status is
not routine even in hospitalized patients whose diseases and
treatment cause Mg-loss. The possibility that long-standing
sub-optimal Mg-intakes might be contributory to disease is
disregarded by the majority of physicians, epidemiologists, and
nutritionists.
It is largely from the early metabolic balance studies that
have come the data indicating that adults might require 6-10
mg/kg/day of Mg, and that infants, children, adolescents, and
pregnant women probably need considerably more. Most of the
relatively recent studies have focused on the least Mg at which
equilibrium can be maintained. The conclusion reached, after
evaluation of metabolic balance studies during the first half of
this century, that metabolic balance can be maintained even in
the presence of deficiency, and that a new equilibrium is
established at intakes more likely to meet needs [143], has not
usually been applied to Mg studies.
One should keep in mind the normal body's ability to limit
Mg-loss and to adapt to marginal intakes for prolonged periods.
For example, rats have shown the capacity to adapt to Mg-intakes
low enough to cause signs of deficiency early, that later
disappear on the same (Mg-deficient) diet, with return of normal
plasma Mg levels [133]. Despite this seeming tolerance of low
dietary Mg, the animals' ability to withstand stress, and their
life spans were reduced as compared with controls on normal
diets. This important study demonstrates the invalidity of
Mg-restriction studies as determinants of Mg-requirements.
Interpreting obligatory urinary Mg-losses by healthy subjects on
diets extremely low in Mg as the amount required [20], or
calculating the minimal amount of dietary Mg that will prevent
hypomagnesemia [199], yield grossly inadequate estimates of
Mg-requirements. Metabolic balance studies at low or marginal
Mg-intakes, that show little or no net loss, might reflect the
normal subjects' capacity to adapt to periods of sub-optimal
Mg-intakes, or (in the case of subjects in secluded, protected
environments) a minimal requirement.
Rather than focusing on how little Mg can be tolerated, it is
preferable to determine how much Mg can be taken without
producing discomfort or adverse reactions. The widespread use of
high dosage Mg as a cathartic, of lower (but still high) dosage
as an antacid, and of pharmacologic Mg-dosage as an
anticonvulsant and anti-hypertensive agent (e. g., in eclampsia),
has provided us with toxicological data as to upper limits of
safety. Unless there is renal immaturity (as in newborn infants)
or renal decompensation, high intakes of Mg have proven safe. In
contrast, Mg-deficiency has been associated with a vast array of
experimental and clinical dysfunctions and diseases of varying
severity. Geographic differences in incidences of diseases with
manifestation in common with some produced by Mg-deficiency in
animals (Reviews: [7, 158, 197, 270, 271, 274]) suggest possible
interactions among host, environmental, and dietary factors that
affect Mg-requirements, retention and distribution. Figure 2
Might the adaptation of weanling rats to sustained low
Mg-intakes, with long-term disadvantages in terms of tolerance to
stress and life expectancy [133] be an experimental analogue of
human sub-optimal Mg intakes from infancy onward, that might be
contributory to many chronic diseases, and even to sudden cardiac
death [268, 270-274, 278]? To what extent do nutritional
imbalances that increase Mg-requirements, and genetic or
physiologic differences, determine whether an individual will
develop disorders against which increasing the Mg-intake might be
protective? We must consider the implications of the early
metabolic balance studies of pregnant women, which show that
women on Mg-intakes in excess of the RDA of 450 mg were in
strongly positive Mg-balance during the third trimester
[141,164], in light of the surveys showing that pregnant
middleclass American women ingest less Mg than half the RDA [14,
155]. To what extent does the developing human fetus suffer from
a deficit that in experimental animals causes malformations,
anemia, growth retardation, and edema [46, 55, 146, 147, 328],
abnormalities depending on the time during gestation, and the
degree of deficiency? Might gestational Mg deficiency (alone and
in combination with other dietary imbalances) contribute both to
maternal complications and to cardiovascular, skeletal and renal
abnormalities, present at birth or becoming manifest later [267,
271, 272, 274]? Do infants born to mothers whose Mg-intakes are
low or suboptimal, especially if their needs are high (as with
teen-aged mothers and those with multiple or frequent births or
who are diabetics, have higher Mg-requirements than can be met by
milk: mother's or formula? This is a matter or urgent concern, in
view of the recent conclusion that, because infant formulas
provide 50-70mg Mg/day, that is the requirement [101, 301].
Breast-fed infants are less subject to hypomagnesemic
hypocalcemia than are formula-fed babies, a possible reflection
of nutrient interactions (high Ca, P, and vitamin D content of
cow's milk) that increase Mg-requirements [271, 272]. How much Mg
is necessary to maintain positive Mg-balances by growing
children? Adolescent boys required 10 to 16 mg/kg/ day to allow
for retention of enough Mg, relative to N, to meet needs for
growth and developments [263]. It may well be that pregnant and
lactating women, athletes-in-training and competition, and
convalescents have comparably high Mg-needs. The Mg provided in
the diet or by supplements to adults in stable physical condition
should be sufficient to allow for the extra needs under
conditions of unexpected stress, and to meet the needs of those
who have higher than average maintenance Mg-requirements. Little
is known of the Mg-needs of those undergoing catabolic processes,
as during chronic disease or in old age. Their low dietary supply
of Mg [312], and the evidence of decreased Mg-absorption [154,
216] suggest that they are likely to be in Mg-deficit. Since
nutrition influences the immunologic response to infection [43],
and Mg plays an important role in humoral and cellular responses
([6, 69, 83, 165, 192, 241] Reviews: [270, 274]), the
Mg-requirements of patients with infections might be elevated.
Differences in plasma and erythrocyte Mg levels in groups with
high predisposition to auto-immune diseases [130] suggest that
they may have different Mg-requirements.
Also requiring study is the effect of dietary imbalances on Mg
requirements. In the developed world, the common imbalances
predisposing to Mg deficiency are predominantly caused by
excesses: of vitamin D, phosphate and calcium, and of fat,
protein and sugar. It is possible that megavitamin or trace
mineral therapy might also adversely affect the retention of Mg.
Metabolic abnormalities in utilization of vitamins and minerals
that interact with Mg, and that might increase its requirements
might also be contributory. Also affecting how much Mg is needed
by the individual is efficacy of absorption and of renal tubular
reabsorption of Mg. Since there are extremes of (Mg)
malabsorption or renal wastage, it seems plausible that there
might be less severe failures of Mg-utilization — whether
genetically determined or produced by disease or dietary factors.
It is quite possible that Mg-needs differ at extremes of altitude
and climate, since plasma and erythrocyte Mg-levels differ [59,
131, 132]. Long-term balance studies are needed to ascertain how
much Mg is needed to maintain equilibrium after the positive
balances produced by supplements that bring the intake up to 6-10
mg/kg/day in stable adults, and after the larger amounts in those
with the greater requirements of anabolism and stress.
Why has Mg been so neglected by so many in the medical
community? Methodological difficulties have hampered accumulation
of baseline data. There is disagreement, even among experts, as
to what normal plasma Mg levels are [5, 319, 320, 326]. Wide
ranges are accepted as normal [309], despite evidence that plasma
Mg is normally maintained within narrow limits, that it is a poor
index of the body Mg-status, and that narrow ranges should be
determined for each laboratory [144, 223, 276, 326]. Renal
Mg-retention of a parenteral Mg-load provides a better clue [35,
37-39, 58, 91, 193, 302, 335], but there are practical drawbacks
to this procedure, except in hospitals. Metabolic balance studies
are important in establishing minimal needs, but this procedure
is far too cumbersome and expensive to be directly applicable to
the individual patient. Mg, being a major intracellular cation,
determination of cellular Mg-levels should provide the best due
as to the adequacy of Mg in the body. Muscle and erythrocyte
Mg-determinations have provided useful data [64, 69, 74, 77,
130-132, 179, 252, 326], but lags before deficiencies are
reflected by changes in these tissues have limited applicability
to a dynamic situation. Mg levels of mixed leukocytes [21, 22,
250] or of lymphocytes [50, 251, 286] might prove to be most
reliable indicator of Mg levels in heart and other internal
organs.
Until we can define, more precisely, how much Mg is needed for
health-maintenance at different ages and under different
physiologic, pathologic, dietary, and environmental conditions,
it is better to provide more than less Mg. For maintenance in the
adult, not less than 6 mg/kg/day for women, and 7-10 mg/kg/day
for men is likely to provide a margin of safety, that will meet
needs even under conditions of increased requirements. For new
tissue formation and repair, higher intakes are desirable,
probably to 15 mg/kg/day and possibly more. Whether increasing
the Mg-intake of humans might reduce the incidence and severity
of diseases with characteristics like those seen in experimental
Mg-deficiency is a provocative possibility.
References
[1] Achari, C., Mishra, K. C., Achari, K., Ramkissun, R.,
Upadhyay, S. N.: J. Indian Med. Assoc.
36: 93-95 (1961).
[2] Aikawa, J. K.: Proc. Soc. Exp.
Biol. Med. 103: 363-366 (1960).
[3] — : Proc. Soc. Exp. Biol.
Med. 104: 461-463 (1960).
[4] Alcantara, E. N.: Ph. D. Thesis,
Univ. of Wisconsin (1970).
[5] Alcock, N. W., MacIntyre, I., Radde,
I.: Nature 205: 89-90
(1965).
[61 Alcock, N. W., Shils, M. E.:
Proc. Soc. Exp. Biol. Med. 145: 855-858
(1974).
[7] Aleksandrowicz, J.: In
Protection of Man's Environment, Pol. Sc. Publ., p. 518-528
(1973).
[8] Am. Heart Assoc.: Circulation
58: 762A-766A (1978).
[9] Am. Med. Assoc. Council on Sci. Aff.: J.
A. M. A. 242: 2335-2338 (1979).
[10] Anderson, T. W., Leriche, W. H., Hewitt, D., Neri, L.
C.: In Cantin, M., Seelig, M. S. (eds).
Magnesium in Health and Disease, SP Medical & Scientific
Books, Jamaica, New York, p. 565-571 (1980). Proc. 2nd Int'l, Mg
Sympos. (Montreal, Canada (1976).
[11] Anderson, T. W., Neri, L. C., Schreiber, G. B.,
Talbot, F. D. F., Zdrojewski, A.: Can. Med.
Assoc. Med. J. 113: 109-113 (1975).
[12] Anghileri, L. J., Heidbreder,
M.: Europ. J. Cancer 13:
291-292 (1977).
[13] Anghileri, L. J., Heidbreder, M., Weiler, G.,
Dermietzel, R.: Exp. Cell Biol.
45: 34-47 (1977).
[14] Ashe, J. R., Scofield, F. A., Gram, M.
R.: Am. J. Clin. Nutr. 32:
28l-286 (1979).
[15] Aurousseau, M., Larvor, P., Durlach,
J.: Proc. 1st Mg Congress. Argentina
(1968).
[16] Bajusz, E.: Nutritional Aspects
of Cardiovascular Diseases. J. B. Lippincott, Philadelphia and
Montreal (1965).
[17] Bajusz, E., Selye, H.: Trans.
N. Y. Acad. Sci. 21: 659-667 (1959).
[18] Bawkin, H.: J. Pediat.
14: 1-10 (1939).
[19] Barker, E. S., Elkinton, J. R., Clark J.
K.: J. Clin. Inv. 38:
1733-1745 (1959).
[20] Barnes, B. A., Cope, O., Harrison,
T.: J. Clin. Inv. 37:
430-440 (1958).
[21] Baron, D. N.: Proc. Roy. Soc.
Med. 62: 945-953 (1969).
[22] Baron, D. N., Ahmed, S. A.:
Clin. Sci. 37: 205-219 (1969).
[23] Baumslag, N., Yeager, D., Levin, L., Petering,, H.
G.: Arch. Env. Health. 29:
186-191 (1974).
[24] Beisel, W. R.: Am. J. Clin.
Nutr. 32: 271-274 (1979).
[25] Bessey, O. A., Adams, D. J. D., Hansen, A.
K.: Pediatrics 20: 33-44
(1957).
[26] Better, O. S., Massry, S. G.:
J. Lab. Clin. Med. 79: 794-800 (1972).
[27] Blaxter, K. L., Wood, W. A.:
Brit. J. Nutr. 6: 144-163 (1952).
[28] Bodansky, M.: Am. J. Clin.
Path. 9: 36-51 (1939).
[29] Bogert, L. J., Trail, R. L.: J.
Biol. Chem. 54: 753-761 (1922).
[30] Boullin, D. J.: J. Physiol.
189: 85-99 (1967).
[31] Brin, M.: Am. K. Clin. Nutr.
24: 704-798 (1971).
[32] Bunce, G. E., Sauberlich, H. E., Reeves, P.
G.: J. Nutr. 86: 406-414
(1965).
[33] Caddell, J. L.: Ann. N. Y.
Acad. Sci. 162: 874-890 (1969).
[34] — : Pediatrics
66: 392-413 (1965).
[35] — : Clin. Pediatr.
14: 449-459, 518-519 (1975).
[36] Caddell, J. L., Olson, R. E.:
J. Pediatr. 83: 124-128 (1973).
[37] Caddell, J. L., Ratananon, N., Trangratapit,
P.: Am. J. Clin. Nutr. 26:
612-615 (1973).
[38] Caddell, J. L., Saier, F. L., Thomason, C.
A.: Am. J. Clin, Nutr. 28:
1099-1104 (1975).
[39] Caddell, J. L., Suskind, R., Sillup, H., Olson, R.
E.: J. Pediatr. 83:
129-135 (1973).
[40] Campbell, B. J., Reinhold, J. G., Cannell, J. J.,
Nourmand, I.: Pahlavi Med. J.
7: 1-17 (1976).
[41] Cantin, M.: Lab. Invest.
22: 558-568 (1970).
[42] Chan, G. M., Tsang, R. C., Chen, I. W., Deluca, H.
F., Steichen, J. J.: J. Pediat.
93: 91-96 (1978).
[43] Chandra, R. K., Newberne, P.
M.: Nutrition, Immunity and Infection.
Plenum Medical Book Co., New York (1977).
[44] Clark. G. M.: Univ. Calif.
Publ. in Physiol. 5: 195-287 (1926).
[45] Cockburn, F., Brown, J. K., Belton, N. K., Forfar, J.
O.: Arch, Dis. Childh. 48:
99-108 (1973).
[46] Cohlan, S. Q., Jansen, V., Dancis, J., Piomelli,
S.: Blood 36: 500-506
(1970).
[47] Consolazio, C. F., Matoush, L. O., Johnson, H. L.,
Nelson, R. A., Krzywick, H. J.: Am. J.
Clin. Nutr. 20: 672-683 (1967).
[48] Coons, C. M., Schiefelbusch, A., Marshall, G. B.,
Coons, R.: Exp. Sta. Bull. # 223 U.S. Dept.
Agric. (1935).
[49] Cooper, J. R., Itokawa, Y., Pincus, J.
H.: Science 164: 74-75
(1969).
[50] Counihan, T. B., Halley, E., Ryan, M. F, Ryan, M.
P.: Irish J. Med. Sci.
147: 327, 331-332 (1978).
[51] Crawford, T., Crawford, M. D.:
Lancet 1: 229-232 (1967).
[52] Cubberley, P. T., Polster, S. A., Schulman, C.
L.: New Engl. J. Med. 272:
628-630 (1965).
[53] Cullumbine, H., Basnayake, V., Lemottee, J.,
Wickramanayake, T. W.: Brit. J. Nutr.
4: 101-111(1950).
[54] Cushard, W. G., Jr., Creditor, M. A., Canterbury, J.
M., Reiss, E.: J. Clin. Endocr. Metab.
34: 767-771 (1972).
[55] Dancis, J., Springer, D., Cohlan, S.
O.: Pediatr. Res. 5: 131
-136 (1971).
[56] Daniels, A. L.: Am. J. Dis.
Child. 62: 568-576 (1941).
[57] Daniels, A. L., Everson, G. J.:
J. Nutr. 11: 327-341 (1936).
[58] Danielson, B. G., Johansson, G., Ljunghall,
S.: In Johansson, G. (ed). Magnesium
Metabolism, Ed. G. Johansson, Pub Almqvist & Wiksell,
Stockholm, Sweden 1979: II: 1-25.
[59] Darlu, P.: Intl. J. Biometeor,
19: 166-173 (1975).
[60] Davis, J. A., Harvey, D. R., Yu, J.
S.: Arch. Dis. Childh, 40:
286-290 (1965).
[61] De Jorge, B. F. B., Delascio, D., Barros de Ulhoa,
Cintra, A., Antunes, M. L.: Obst. Gynec.
25: 253-254 (1965).
[62] Douglas, W. W., Rubin, R. P.:
J. Physiol. 175: 231-241 (1964).
[63] Draper, H. H.: J. Nutr.
83: 65-72(1964).
[64] Drenick., E. J., Brickman, A.
S.: In Durlach, J. (ed.) Proc. 1st Int'l.
Sympos. on Mg, Vittel, France, 1971, p. 491-498 (1973).
[65] Drenick, E. J., Hunt, I. F., Swendseind, M.
E.: Clin, Endocr. Metab.
29: 1341-1348 (1969).
[66] Drews, L. M., Kies, C., Fox, H.
M.: Am. J. Clin. Nutr. 32:
1893-1897 (1979).
[67] Duckworth, J., Warnock, G.M.:
Nutr. Abstr. Rev. 12: 167-183 (1942).
[68] Dull, T.: Clin. Res.
11: 404-405 (1963).
[69] Durlach, J.: In Cantin, M.,
& M. S. Seelig, Magnesium in Health and Disease, SP Medical
& Scientific Books, Jamaica, New York, p. 883-909 (1980).
Proc. 2nd Intl. Mg Sympos., Montreal, Canada (1976).
[70] — : Vie Med.
41: 966-969 (1960).
[71] — : Nouv. Arch. Hosp.
42: 31-40 (1970).
[72] — : Diabete
19: 99-113 (1971).
[73] — : In: Vinken, P. J.
Bruyn, G. W. (eds), Handbook of Clinical Neurology 28,
North-Holland Publ. Co., Amsterdam-Oxford-New York, p. 545-579
(1976).
[74] — : Spasmophilia and
Magnesium Deficit. Masson Co., Paris, France, p. 63-136
(1969).
[75] Durlach, J., Lebrun, R.: Ann.
Endocr. (Paris) 21: 244-252 (1960).
[76] Durlach, J., Rayssiguier, Y., Laguitton,
A.: Med. Nutr. 16: 15-21
(1980).
[77] Dyckner, T., Wester, P. O.: In:
Cantin, M., Seelig, M. S., Magnesium in Health and
Disease, SP Medical & Scientific Books, Jamaica, New York, p.
551-557 (1980). Proc. 2nd Intl. Mg Sympos. Montreal, Canada
(1976).
[78] Ebadi, M. S., Bostad, R., Pellegrino, R.
J.: J. Neur., Neurosurg., Psychiatr.
32: 393-398 (1969).
[79] Ebel, H., Gunther, T.: J, Clin.
Chem. Clin. Biochem. 18: 257-270 (1980).
[80] Ebels, E. J., Blokzijl, F. J., Troelstra, J.
A.: Helv. Paed. 3: 310-324
(1965).
[81] Elin, R. J.: Proc. Soc. Exp.
Biol. Med. 142: 1159-1161 (1973).
[82] — : In: Cantin, M.,
Seelig, M. S., Magnesium in Health and Disease. SP Medical
& Scientific Books, Jamaica, New York, p. 113-124 (1980).
Proc 2nd Intl. Mg Sympos. Montreal, Canada (1976).
[83] — : Proc. Soc. Exp. Biol.
Med. 148: 620-624 (1975).
[84] Elin, R. J., Tan, H. K.: Blood
49: 657-664 (1977).
[85] Ertel, N. H., Reiss, J. S., Spergel,
G.: New Engl. J. Med. 280:
260-262 (1969).
[86] Feenstra, L., Wilkens, J. H.:
Ned. T. Geneesk 109: 615-19 (1965).
[87] Fehlinger, R.: Rev. Franc.
Endocr. Clin. Nutr. Metab. 20: 501-513
(1979).
[88] — : Magnesium Bull.
2: 40-47 (1980).
[89] Feigin, L, Wolf A.: J. Pediatr.
45: 243-263 (1954).
[90] Fenn, W. O., Goetisch, M.: J.
Biol. Chem. 120: 41-51 (1937).
[91] Fitzgerald, M. G., Fourman, P.:
Clin. Sci. 15: 635-647 (1956).
[92] Flink., E. B.: In: Cantin,
M., Seelig, M. S.: Magnesium in Health and
Disease, SP Medical & Scientific Books, Jamaica, New York, p.
865-882 (1980). Proc. 2nd Intl. Mg Sympos. Montreal, Canada
(1976).
[93] Flink., E. B., Flink., P. F., Shane, S. R., Jones, J.
E., Steffes, P. E.: Clin. Res.
21: 884 (1973).
[94] Flink, E. B., McCollister, R., Prasad, A. S., Melby,
J. C., Doe, R. P.: Ann. Intern. Med.
47: 956-968 (1957).
[95] Flink, E. B., Morano, G. D., Morabito, R. A., Shane,
S. R., Scobbo, R. R.: In: Cantin, M.,
Seelig, M. S. (eds). Magnesium in Health and Disease, SP
Medical & Scientific Books, Jamaica, New York, p. 73-78
(1980). Proc. 2nd Intl. Mg Sympos., Montreal, Canada (1976).
[96] Flink. E. B., Shane, S. R., Scobbo, R. R.,
Blehschmidt, N. G., McDowell, P.:
Metabolism 28: 858-865 (1979).
[97] Flink. E. B., Stutzman, F. L., Anderson, A. R.,
Konig,, T., Fraser, R.: J. Lab, Clin. Med.
43: 169-183 (1954).
[98] Flowers, C. E. Jr.: Am, J.
Obst. Gyn. 91: 763-776 (1965).
[99] Fodor, J. G., Pfeiffer, C. J., Papezik, V.
S.: Can. Med. Assoc. J.
108: 1369-1373 (1973).
[100] Food & Nutr. Board Nat'l Res. Council: Publ. Natl.
Acad. Sci. Washington, D. C. Intl. Standard # 0-309-00377-3
(1980).
[101] Food & Nutr. Board, Nat'l Res. Council: Recommended
Dietary Allowances. Ed. 9, Natl, Acad. Sci., Washington, D.C.
(1980).
[102] Forbes, R. M.: J. Nutr.
80: 321-326 (1963).
[103] Fourman, P., Morgan, D. B.:
Proc. Nutr. Soc. 21: 34-41 (1962).
[104] Freeman, R. M., Pearson, E.:
Am. J. Med. 41: 645-656 (1966).
[105] Friderichsen, C.: Lancet
1/6: 85-86 (1939).
[106] Frimpter, G. W., Andelman, R., George, W.
F.: Am. J. Clin. Nutr. 22:
794-803 (1969).
[107] Garnett, E. S., Barnard, D. L., Ford, J., Goodbody,
R. A., Woodehouse, M. A.: Lancet
1: 914-916 (1969).
[108] Gershoff S. N.: Proc. Soc.
Exp. Biol. Med. 127: 1207-1210 (1968).
[109] Gershoff S. N., Prien, E. L.:
Am. J. Clin. Nutr. 20: 393-399, 1967.
[110] Gitelman, H. J., Graham, J. B., Welt, L.
G.: Ann. N. Y. Acad. Sci.
162: 856-864 (1969).
[111] Gittleman, I. F., Pincus, J. B., Schmertzler,
E.: Am. J. Dis Child. 107:
119-124 (1964).
[112] Goldsmith, L.A.: J. Nutr.
93: 87-102 (1967).
[113] Gormican, A., Catli, E.: Nutr.
Metab. 13: 364-377 (1971).
[114] Greenberg, D. M.: Biochem.
8: 269-300, 1939.
[115] Greenberg, D. M., Tufts, E.
V.: Am. J. Physiol. 121:
416-423 (1938).
[116] Greger, J. L., Bennett, O. A., Abernathy, R.
P.: Fed. Proc. 36: 1130
(1977).
[117] Greger, J. L., Baligar, R., Abernathy, R. P.,
Bennett, O. A., Peterson, T.: Am. J. Clin.
Nutr. 31: 117-121 (1978).
[118] Greger, J. L., Gruner, S. M., Ethyre, G.M.,
Abernathy, R. P., Sickles, V.: Nutr. Rep.
Int'l. 20: 235-243 (1979).
[119] Greger, J. L., Huffman, J., Abernathy, R. P.,
Bennett, O. A., Resneck, S. E.: J. Food Sc.
44: 1765, 1766. 1771 (1979).
[120] Haijamae, H., MacDowell, I.
G.: Acta Paediatr. Scand.
61: 591-596 (1972).
[121] Hall, D. G.: Obst. Gyn.
9: 158-162 (1957).
[122] Hambidge, K. M., Walravens, P.
A.: In: Prasad, A. S., Trace
Elements in Human Health and Disease 1, Academic Press. New York,
p. 21-32 (1976).
[123] Harrington, D. D.: Brit. J.
Nutr. 34: 45-57 (1975).
[124] Hartenstein, H., Gardner, L.
I.: New Engl. J. Med. 274:
266-268 (1966).
[125] Hathaway, M. L.: Home Econ.
Res. Report # 19, Agric. Res. Serv. U.S.D.A. Washington, D.C.
(1962).
[126] Haury, V. G., Cantarow, A.: J.
Lab. Clin. Med. 27: 616-622 (1942).
[127] Haywood, J., Selvester, R.:
Clin. Med. 10: 87 (1962).
[128] Heaton, F. W.: In Cantin,
M., & Seelig, M. S., Magnesium in Health and Disease, SP
Medical & Scientific Books, Jamaica, New York, 43-55 (1980).
Proc. 2nd Int'l. Mg Sympos. Montreal, Canada (1976).
[129] Hellerstein, E. E., Nakamura, M., Hegsted, D. M.,
Vitale, J.J.: J. Nutr. 71:
339-346 (1960).
[130] Henrotte, J. G.: C. R. Acad.
Sc. (Paris) 285: 737-739 (1977).
[131] Henrotte, J. G., Benech, A., Pineau,
M.: In: Cantin, M., Seelig, M. S.
Magnesium in Health and Disease, SP Medical & Scientific
Books, Jamaica, New York, p. 929-934 (1980). Proc. 2nd Int'l Mg
Sympos., Montreal, Canada (1976).
[132] Henrotte, J. G., Pechery, C., Durlach, J., De
Traverse, P.M.: In: Durlach, J.
(ed.) Proc. 1st Int'l Mg Sympos. 1971, Vittel, France 2, p.
573-576 (1973).
[133] Heroux, O., Peter, D., Heggtveit,
A.: J. Nutr. 107:
1640-1652 (1977).
[134] Hills A. G., Parsons, D. W., Webster, G. D.,
Rosenthal, O.: J. Clin. Endocr. Metab.
19: 1192-1211(1959).
[135] Hodgkinson, A., Heaton, F. W.:
Clin. Chim. Acta 11: 354-362 (1965).
[136] Hoffstrom, K A,: Skand. Arch.
Physiol. 23: 326-420 (1916).
[137] Holtmeier, H. J.: In:
Heilmeyer, L. (ed.) Ernährungswissenschaften, V:
Sympos. Freiburg, 1967, George Thieme Verlag, Stuttgart, W.
Germany, p. 111-152 (1968).
[138] — : In: Durlach,
J. (ed.) Proc. 1st Int'l Mg Sympos. 1971, Vittel, France 1,
p. 397-417 (1971).
[139] Hsu, J. M.: Proc. Soc. Exp.
Biol. Med. 119: 177 -180 (1965).
[140] Hummel, F. C., Hunscher, H. A., Bates, M. F.,
Bonner, P., Macy, I. G.: J. Nutr.
13: 263-278 (1937).
[141] Hummel, F. C., Sternberger, H. R., Hunscher, H. A.,
Macy, I. G.: J. Nutr. 11:
235-255 (1936).
[142] Hungerford, G. F., Bernick.,
S.: In: Cantin, M., Seelig, M. S.,
Magnesium in Health and Disease, SP Medical & Scientific
Books, Jamaica, New York, p. 659-670 (1980). Proc. 2nd Int'l. Mg
Sympos., Montreal, Canada (1976).
[143] Hunscher, H. A.: J. Am. Diet.
Assoc. 39: 209-219 (1961).
[144] Hunt, B. J.: Clin. Chem,
15: 979-996 (1969).
[145] Hunt, S. M., Schofield, F A.:
Am. J. Clin. Nutr. 22: 367-373 (1969).
[146] Hurley, L. S.: In:
Durlach, J. (ed.) Proc. 1st Int'l Mg Sympos., Vittel,
France, p. 481-492 (1971).
[147] Hurley, L. S., Cosens, G., Therrault, L.
L.: J. Nutr. 106:
1254-1260 (1976).
[148] Irwin, M. I., Feeley, R. M.:
Am. J. Clin. Nutr. 20: 816-824 (1967).
[149] Irwin, M. I. & Wiese, H.
F.: J. Nutr. 74: 217-225
(1961).
[150] Itokawa, Y., Inoue, K., Natori, Y., Okazaki, K.,
Fujiwara, M.: J. Vitaminol.
18: 159-164 (1972).
[151] Itokawa, Y., Tanaka, C., Kimura,
M.: Metabolism 21: 371-375
(1972).
[152] Itokawa, Y., Tseng, L. F., Fujiwara,
M.: J. Nutr. Sci. Vitaminol.
20: 249-255 (1974).
[153] Jabir, F. K., Roberts, S.D., Womersley, R.
A.: Clin. Sci. 16: 119-124
(1957).
[154] Johansson, G.: Scand. J. Urol.
Nephrol. Suppl. 51: 1-16 (1979).
[155] Johnson, N. E., Phillips, C.:
In: Cantin, M., Seelig, M. S., Magnesium in Health and
Disease, SP Medical & Scientific Books, Jamaica, New York, p.
827-831 (1980). Proc. 2nd Int'l Mg Sympos. Montreal, Canada,
1976.
[156] Jones, J. E., Shane, S. R., Jacobs, W. H., Flink, E.
B.: Ann. N.Y. Acad. Sci.
162: 934-946 (1969).
[157] Kalbfleisch, J. M., Lindeman, R. D., Smith, W,
O.: J. Lab. Clin. Med. 48:
833-834 (1961).
[158] Karppanen, H., Pennanen, R., Passinen,
L.: Adv. Cardiol. 25: 9-24
(1978).
[159] Keating, F. R., Jr., Jones, J. D., Elveback, L. R.,
Randall, R. T.: J. Lab. Clin. Med.
73: 825-834 (1969).
[160] Kelsay, J. L., Behall, K. M., Prather, E.
S.: Am. J. Clin. Nutr. 32:
1876-1880 (1979).
[161] Knochel, J. P.: Arch. Int.
Med. 137: 203-220 (1977).
[162] Korbitz, B. C.: J. Vitaminol.
16: 137-139 (1970).
[163] Landes, R. R., Melnick. I., Sierakowski, R.,
Finlayson, B.: In Seelig, M. S. (ed),
Nutritional Imbalances in Infant and Adult Diseases, SP Medical
& Scientific Books, Jamaica, New York, p. 9-21 (1977).
[164] Landsberg., E.: Zschr.
Geburts. Gyn. 75: 53-98 (1914).
[165] Larvor, P.: In Cantin, M
&Seelig, M. S., Magnesium in Health and Disease, SP
Medical & Scientific Books, Jamaica, New York, p. 201-224
(1980). Proc. 2nd Int'l Mg Sympos., 1976, Montreal, Canada.
[166] Larvor, P., Durlach, J.: Vie
Med. 35: 4359-4368 (1973).
[167] Lazard, E. M.: Am. J. Obst.
Gyn. 9: 178-188 (1925).
[168] Lebenthal, E.: In:
Lifshitz, F. (ed.) Clinical Disorders in Pediatric
Nutrition, Marcel Dekker Press, New York (in press, 1981).
[169] Lehr, D.: In Bajusz, E. (ed.)
Electrolytes and Cardiovascular Diseases 1, S. Karger, Basel/New
York, p. 248-273 (1965).
[170] — : Ann. N.Y. Acad. Sci.
156: 344-378 (1969).
[171] Lehr, D., Chau, R., Kaplan,
J,: In Bajusz, E. & Rona, G. (eds.).
Recent Advances in Cardiac structure and Metabolism, I.
Myocardiology, University Park Press, Baltimore, p. 684-698
(1972).
[172] Leitch I., Hepburn, A.: Nutr.
Abstr. Rev. 31: 389-401 (1961).
[173] Lemann, J., Lennon, E. J., Piering, W. K., Prien, E.
L., Ricanati, E. S.: J. Lab. Clin. Med.
75: 578-585 (1970).
[174] Lemann, J.; Piering, W. F., Lennon, E.
J.: J. Lab. Clin. Med. 70:
987-988 (1967).
[175] Lennon, B. J.: In Seelig,
M. S. (ed), Nutritional Imbalances in Infant and Adult
Diseases, SP Medical & Scientific books, Jamaica, New York,
p. 127-139 (1977).
[176] Lennon, E. J., Lemann, J, Jr., Piering, W. F.,
Larson, L. S.: J. Clin. Inv.
53: 1424-1433 (1974).
[177] Leveille, G. A.: Dietary
fiber. Food & Nutr. News 47: (1976).
[178] Lieber, I. I.: Prensa Med.
Argentina 48: 44-52 (1961).
[179] Lim, P., Jacob, E., Dong, S., Khoo, O.
T.: J. Clin. Path. 22:
417-421 (1969).
[180] Lindeman, R. D.: In:
Cantin, M., Seelig, M. S., (eds.). Magnesium in Health
and Disease, SP Medical & Scientific Books, Jamaica, New
York, p. 381-399 (1980). Proc. 2nd Int'l Mg Sympos., 1976,
Montreal, Canada,
[181] Lindeman, R. D., Adler, S., Yiengst, M. J., Beard,
E. S.: J. Lab. Clin. Med.
70: 236-245 (1967).
[182] Linden, V.: In Seelig, M.
S. (ed.), Nutritional Imbalances in Infant and Adult
Diseases, SP Medical & Scientific Books, Jamaica, New York,
p. 23-42 (1977).
[183] Linden, V.: Brit. Med. J.
3: 647-750 (1974).
[184] Lowe, C. U.: Am. J. Clin.
Nutr. 25: 245-254 (1972).
[185] Lutwak, L., Singer, F. R., Urist, M.
R.: Ann. Intern. Med. 80:
630-644 (1974).
[186] McCance, R. A., Widdowson, E.
M.: J. Physiol. 101: 44-85
(1942).
[187] —, — : J. Physiol.
101: 304-313 (1942).
[188] McCance, R. A., Widdowson, E. M., Lehmann,
H.: Biochem. J. 36:
686-691 (1942).
[189] McCance, R. A., Widdowson, E.
M.: Brit. Med. Bull, 17:
132-136 (1961).
[190] McCollister, R. J., Flink, E. B., Lewis, M.
D.: J. Clin. Nutr. 12:
415-420 (1963).
[191] McCormick, D. B., Gregory, M. E., Snell, E.
E.: J. Biol. Chem. 236:
2076-2084 (1961).
[192] McCoy, J. H., Kenney, M. A.:
J. Nutr. 105: 791-797 (1975).
[193] McDonald, J. T., Margen, S.:
Am. J. Clin. Nutr. 32: 823-833 (1979).
[194] McHale, M., Kies, C., Fox, H.
M.: J. Food Sci. 44:
1412-1417 (1979).
[195] McKey, B. V., LaFont, F. M., Borchers, G. L.,
Navarrete, D. A., Holmes, J. O.: Fed. Proc.
21: 310 (1962).
[196] Macy, I. G.: Nutrition &
Chemical Growth in Childhood, III. Charles C. Thomas,
Springfield, III, 1951.
[197] Marier, J, K.: Rev. Can. Biol.
37: 115-125 (1978).
[198] Marsh, A. G., Ford, D. C., Christensen, D.
K.: J. Am. Diet. Assoc.
51: 441-446 (1967).
[199] Marshall, D. H., Nordin, B. E. C., Speed,
R.: Proc. Nutr. Soc. 35:
163-173 (1976).
[200] Marshall, M. W., Iacono, J. M., Young, C. W.,
Washington, V. A., Slover, H. T., Leapley, P.
M.: J. Am Diet. Assoc. 66:
470-481 (1975).
[201] Martin, H. E.: Ann. N. Y.
Acad. Sci. 162: 891-900 (1969).
[202] Martin, J. E., Jones, R.: Am.
Heart J. 62: 206-210 (1961).
[203] Maynard, L.A., Boggs, D., Fisk, G., Seguin,
D.: J. Nutr. 64: 85-97
(1958).
[204] Mays, E. T., Randall, H. T., Deweese, B.
M.: Surgery 67: 780-788
(1970).
[205] Mellinghoff K.: Dtsch. Arch.
Klin. Med. 195: 475-480 (1949).
[206] Mellinghoff K., An Lessen, W.:
Dtsch. Arch. Klin. Med. 194: 285-293 (1949).
[207] Melnick, L, Landes, R. R., Hoffman, A.
A.: In Durlach, J. (ed.) Proc. 1st
Int'l Mg Sympos., Vittel, France, 1971, 2, p. 69-72 (1973).
[208] Melnick, L, Landes, R. R., Hoffman, A. A., Burch, J.
F.: J. Urol. 105: 119-122
(1971).
[209] Michelis, M. F., Drash, A. L., Linarelli, L. G.,
Derubertis, F. R., Davis, B. B.: Metabolism
21: 905-920 (1972).
[210] Michiel, R. R., Sneider, J. S., Dickstein, R. A.,
Hayman, H., Eich, R. H.: New Engl. J. Med.
198: 1005-1007 (1978).
[211] Miller, R. T., Faloon, W. W., Lloyd, C.
W.: J. Clin. Endocr. Metab.
18: 1178-1185 (1958).
[212] Mishra, K.: Rev. Can. Biol.
19: 143-153 (1960).
[213] Monteleone, .J. A., Lee, J. B., Tashjian, A. H.,
Cantor, H. E.: Ann. Intern. Med.
82: 670-672 (1975).
[214] Morris, E. R., O'Dell, B. L.:
J. Nutr. 75: 77-85 (1961).
[215] —, — : J. Nutr.
81: 175-181 (1963).
[216] Mountokalakis, Th., Singhellakis, P. N., Alevizaki,
C. C., Caramanakos, E., Ikkos, D. G.: Rev.
Franc. Endocr. Clin. Nutr. Metab. 17: 229-232
(1976).
[217] Munoz, J. M., Sandstead, H. H., Jacob, R. A., Logan,
G. M. Jr., Reck, S. J., Klevay, L. M., Dintzis, F. R., Inglett,
G. E., Shuey, W. C.: Am. J. Clin. Nutr.
32: 580-592 (1979).
[218] Nabarro, D. N., Spencer, A. G., Stowers, J.
M.: Quart. J. Med. 21:
225-243 (1952).
[219] Neri, L. C., Hewitt, D., Schreiber, G. B., Anderson,
T. W., Mandel, J. S., Zdrojewsky, A.: J. A.
W. W. A. 67: 403-409 (1975).
[220] Neri, L. C., Marier, J. K.:
In: Naito, H. K., Gerrity, R. G. (eds), Nutrition and
Cardiovascular Diseases, SP Medical & Scientific Books,
Jamaica, New York (in press.).
[221] Newman, R. L.: Obst. Gyn.
10: 51-55 (1957).
[222] Nordio, S., Donath, A., Macagno, F., Gatti,
R.: Acta Paediat. Scand.
60: 441-448, 449-455 (1971).
[223] Nuoranne, P.: Nord. Vet. Med.
30: 71-73 (1978).
[224] Nuoranne, P., Raunio, R. P., Saukko, P., Karppanen,
H.: Brit. J. Nutr. 44:
53-60 (1980).
[225] O'Dell, B. L., Morris, E. R., Regan, W.
O.: J. Nutr. 70: 103-110
(1960).
[226] Parsons, R. S.: Med. J.
Austral. 1: 883 -884 (1958).
[227] Paunier, L.: Med. Nutr.
16: 31-36 (1980).
[228] Paunier, L., Radde, I. C., Kooh, S.
W.: Pediatrics 41: 385-402
(1968).
[229] Paunier, L., Sizonenko, P. C.:
J. Pediatr. 88: 51-55 (1976).
[230] Petersen, V. P.: Acta Med.
Scand. 173: 285-298 (1963).
[231] Petrunkina A.: Ztschr.
Kinderheilk. 56: 219-226 (1934).
[232] Prasad, A. S.: Zinc in Human
Nutrition, CRC Press, Boca Raton, Fl. 1979.
[233] Prein, E. L.: J. A. M. A.
19: 177 (1965).
[234] Pritchard, J. A., Stone, S.
R.: Am. J. Obst. Gynec.
99: 752-765 (1967).
[235] Raab, W.: In Bajusz, E.
& Rona, G. (eds.) Recent Advances in Studies on Cardiac
Structure and Metabolism, I. Myocardiology, University Park
Press, Baltimore, p. 707-713 (1972).
[236] Raab, W.: Ann. N. Y. Acad.
Sci. 147: 627-686 (1969).
[237] Rapado, A., Castrillo, J. M.:
In: Cantin, M., Seelig, M. S. (eds.) Magnesium in Health
and Disease, SP Medical & Scientific Books, Jamaica, N.Y., p.
355-364 (1980). Proc., 2nd Intl Mg Symposium, 1976, Montreal,
Canada.
[238] Rapado, A., Castrillo, J. M.:
Ibid, p. 485-497 (1980).
[239] Rayssiguier, Y.: Horm. Metab.
Res. 9: 309-314 (1977).
[240] Rayssiguier, Y., Gueux, E.:
Ann. Nutr. Aliment. 33: 545-546 (1979).
[241] Rayssiguier, Y., Larvor, P., Augusti, Y., Durlach,
J.: Ann. Biol. Anim. Bioch. Biophys.
17: 147-152 (1977).
[242] Rayssiguier, Y., Larvor, P.:
In: Cantin, M., Seelig, M. S. (eds.) Magnesium in Health
and Disease, SP Medical & Scientific Books, Jamaica, N. Y.,
p. 67-72 (1980), Proc. 2nd Intl Mg Sympos. 1976, Montreal,
Canada.
[243] Reinhold, J. G., Bahram, F., Parichehr, A.,
Ismall-Beigi, F.: J. Nutr.
106: 493-503 (1976).
[244] Rigo, J.: In: Durlach,
J. (ed.) Proc. 1st Intl Mg Sympos., Vittel, France, p.
214-228 (1971).
[245] Rigo, J.: Gerontologia
2: 25-33(1965).
[246] Rigo, J., Li, B. N., Zelles, T., Szelenyi, I., Sos,
J.: Acta Physiol. Acad. Sci. Hung. Suppl.
25: 40 (1965).
[247] Rigo, J., Simon, C., Hegyvari, C., Sos,
J.: Acta Med. Acad. Sci. Hung.
19: 231-236 (1963).
[248] Rigo, J., Szelenyi, I., Sos,
J.: Acta Physiol. Acad. Sc. Hung.
32: (Suppl.) 16 (1967).
[249] Rosenbloom, A.: Metabolism
26: 1033-1039 (1977).
[250] Ross, R. S., Seelig, M. S., Berger, A.
R.: In: Cantin, M., Seelig, M. S.
(eds.). Magnesium in Health and Disease, SP Medical &
Scientific Books, Jamaica, New York, p. 7-15 (1980). Proc. 2nd
Intl. Mg Sympos., 1976, Montreal, Canada.
[251] Ross, R. S., Seelig, M. S., Berger, A.
R.: (this Symposium).
[252] Rousselet, F., Durlach, J.:
In: Durlach, J., Proc. 1st Int'l Mg Sympos., Vittel,
France, p. 65-90 (1971)
[253] Rubin, H.: Proc. Nat. Acad.
Sci. USA 72: 3551-3555 (1975).
[254] Rubin, R. P.: Pharmacol. Rev.
22: 389-428 (1970).
[255] Runeberg, L., Collan, Y., Jokinen, E. J.,
Lahdevirta, J., Aro, A.: Am. J. Med.
59: 873-991 (1975).
[256] Salet, J., Polonovski, C., de Gouyon, F., Pean, G.,
Melekian, B., Fournet, J. P.: Arch. Franc.
Pediat. 23: 749-768 (1966).
[257] Salet, J., Polonovski, C., Fournet, de Gouyon, F.,
Aymard, P., Pean, G., Taillemite, .J. L.:
Arch. Franc. Pediat. 27: 550-551 (1970).
[258] Sandstead, H. H., Klevay, L. M., Jacob, R. A.,
Munoz, J. M., Logan, G. M., Jr., Reck., S.
J.: In: Inglett, G. E., Falkenhag, S.
I. (eds.) Dietary Fibers: Chemistry and Nutrition, Academic
Press, New York, p. 147-156 (1979).
[259] Savoie, L. L.: Path. Biol.
20: 19-20; 751-756 (1972).
[260] Savoie, L. L., Delorme, B.: In
Cantin, M. & Seelig, M. S., Magnesium in Health
& Disease. SP Medical & Scientific Books, Jamaica, New
York, p. 537-544 (1980). Proc. 2nd Int'l. Mg Sympos. 1976,
Montreal, Canada.
[261] Sawyer, M., Baumann, L., Stevens,
F.: J. Biol. Chem. 33:
103-109 (1918).
[262] Schlutz, F. W., Morse, M., Oldham,
H.: Am. J. Dis. Child. 46:
757-774 (1933).
[263] Schwartz, R., Walker, G., Linz, M. D., MacKellar,
I.: Am. J. Clin. Nutr. 26:
510-518 (1973).
[264] Schwartz, R., Woodcock, N. A., Blakeley, J. D.,
MacKellar, I.: Am. J. Clin. Nutr.
26: 519-523 (1973).
[265] Schwarz, R,: In: Harris,
R. S., Wool, I. G. (eds.) Vitamins and Hormones 20, Academic
Press, New York, p. 463-484 (1962).
[266] Scriver, C. R.: Pediatrics
26: 62-74 (1960).
[267] Seelig, M. S.: In: Lavery,
P. C. (ed.) Nutrition in Pregnancy (in press, 1981).
[268] — : In: Naito, H.
K., Gerrity, R. (eds.) Nutrition and Cardiovascular
Diseases, SP Medical & Scientific Books, Jamaica, New York
(in press, 1981).
[269] — : In: Durlach,
J. (ed.), Proc. 1st Int'l Mg Sympos., Vittel, France, p.
11-38 (1971).
[270] — : Biol. Tr. Elem. Res.
1: 273-297 (1979).
[271] — : Magnesium Deficiency
in the Pathogenesis of Disease. Early Roots of Cardiovascular,
Skeletal, and Renal Abnormalities. Ser. Ed., L. V.
Avioli, Plenum Medical Books, New York (1980).
[272] — : Cardiovasc. Med.
3: 637-650 (1978).
[273] — : In: Bajusz, E.,
Rona, G. (eds.) Recent Advances in Studies on Cardiac
Structure and Metabolism. I. Myocardiology, University Park
Press, Baltimore, p. 615-625; 626-638 (1972).
[274] — : In: Lifshitz,
F. (ed.) Clinical Disorders in Pediatric Nutrition, Marcel
Dekker, New York (in press 1981).
[275] — : Am. J. Clin.
Nutrition, 14: 342-390 (1964).
[276] Seelig, M. S., Berger, A. K,:
New Eng J. Med. 290: 974-975 (1974).
[277] Seelig, M. S., Berger, A. K., Spielholz,
N.: Dis. Nerv. Syst. 36:
461-465 (1975).
[278] Seelig, M. S., Haddy, F. J.:
In: Cantin, M., Seelig, M. S. (eds.). Magnesium in
Health and Disease, SP Medical & Scientific Books, Jamaica,
NY, 605-638 (1980). Proc. 2nd Int'l Mg. Sympos., 1976, Montreal,
Canada.
[279] Seelig, M. S., Heggtveit, H.
A.: Am. J. Clin. Nutr. 27:
59-79 (1974).
[280] Seelig, M. S., Vitale, J. J.:
In: Durlach, J. (ed.) Proc. 1st Intl. Mg Sympos., 1971,
Vittel, France, p. 515-522 (1973).
[281] Selye, H.: Acta Endocr.
28: 273-278 (1958).
[282] — : Am. Heart J.
55: 805-809 (1958).
[283] — : The Chemical
Prevention of Cardiac Necroses, The Ronald Press Co., New York
(1958).
[284] — : The Pluricausal
Cardiomyopathies, Charles C. Thomas, Springfield, Illinois
(1961).
[285] Shukers, C. F., Knott, E. M., Schultz, F.
W.: J. Nutr. 22: 53-64
(1941).
[286] Silver, B. B., Seelig, M. S.:
(this Symposium).
[287] Slater, J. E.: Brit. J. Nutr.
15: 83-97 (1961).
[288] Slavin, J. L., Marlett, J. A.:
Am, J. Clin. Nutr. 33: 1932-1939 (1980).
[289] Sos, J.: In: Bajusz,
E. (ed.) Electrolyte and Cardiovascular Diseases, S. Karger,
Basel/New York; Williams & Wilkins, Baltimore, p. 161-180
(1965).
[290] Spencer, H., Lesniak, M., Kramer, L., Osis,
D,: In: Cantin, M., Seelig, M. S.,
(eds.) Magnesium in Health and Disease, SP Medical &
Scientific Books, Jamaica, New York, p. (1980). Proc. 2nd Int'l
Mg Sympos., 1976, Montreal, Canada.
[291] Spencer, I. O. B.: Lancet
1: 1288-1290 (1968).
[292] Srivastava, U. S., Nadeau, M. J., Guennou,
L.: Nutr. Rep. Intl. 18:
235-242 (1978).
[293] Steiner, A. J.: In: Naito,
H. K., Gerrity, R. (eds.) Nutrition and Cardiovascular
Diseases, SP Medical and Scientific Books, Jamaica, New York, (in
press, 1981). Proc. 19th Ann. Mtg. of Am. Coll. of Nutr,
Minneapolis, 1978.
[294] Stephenson, M. G., Butler, L. C., Adams, Y.
L.: Fed. Proc. 29: 696
(1970).
[295] Stewart, A., K., Magee, A. C.:
J. Nutr. 82: 287-295 (1964).
[296] Stolley, P. D.: Ann. Intern.
Med. 76: 661-663 (1972).
[297] Stromme, J. H., Nesbakken, R., Normann, T.,
Skjortem, F., Skyberg, D., Johannessen, B.:
Acta Paediat, Scand. 58: 433-444 (1969).
[298] Sunderman, F. W.: AM J. Clin.
Path. 17: 169-180 (1947).
[299] Swanson, W. W,: Am. J. Dis.
Child. 43: 10-18 (1932).
[300] Szelenyi, I.: In: Durlach,
J. (ed). Proc. 1st Int'l Mg Sympos., Vittel, France, p.
195-211 (1971).
[301] — : World Rev. Nutr.
Diet 17: 189-224 (1973).
[302] Thoren, L.: Acta Chir. Scand.
(Suppl. 306): 1-65 (1963).
[303] Tipton, I. H., Stewart, P. L.:
Dev. Appl. Spectroscopy 8: 40-50 (1970).
[304] Touitou, Y., Touitou, C., Bogdan, A., Beck, H.,
Reinberg, A.: Clin. Chim. Acta
87: 35-41 (1978).
[305] Tower, D. B.: Am. J. Clin.
Nutr. 4: 329-344 (1954).
[306] Turner, T. L., Cockburn, F., Forfar, J.
O.: Lancet 1: 283-284
(1977).
[307] U.S. Dept. Agric. Nationwide Food Consumption
Survey, 1977-1978. Preliminary Report $2. U.S.D.A. Sci.
& Educ. Adm. (1980).
[308] U. S. Senate Select. Comm. on Nutr. & Human
Needs, Dietary Goals for the U.S. U.S. Govt. Printing
Office, Washington, D.C., 1978.
[309] Unsigned. New Engl. J. Med.
290: 39-49 (1974).
[310] Vallee, B. L. L.: In: The
Enzymes, P. Boyer (ed), Academic Press, N. Y., p.
225-270 (1960).
[311] Varo, P.: Int'l. J. Vitamin
Nutr. Res. 44: 267-273 (1974).
[312] Vir, S. C., Love, A. H. G.:
Am. J. Clin. Nutr. 32: 1934-1947 (1979).
[313] Vitale, J. J., Hellerstein, E. E., Hegsted, D. M.,
Nakamura, M., Farbman, A.: Am. J. Clin.
Nutr. 7: 13 -22 (1959).
[314] Vitale, J. J., Velez, H., Guzman, C., Correa,
P.: Circ. Res. 12: 642-650
(1963).
[315] Vitale, J, J., White, P. L., Nakamura, M., Hegsted,
D. M., Zamcheck, N., Hellerstein, E. E.: J.
Exp. Med. 106: 757-766 (1957).
[316] Wachstein, M., Graffeo, L. W.:
Obst. Gyn. 8: 177-180 (1956).
[317] Wachstein, M., Gudaitis, A.:
J. Lab. Clin. Med. 40: 550-557 (1952).
[318] Wacker, W. E. C.: Magnesium
and Man. Harvard Univ. Press., Cambridge, Mass. (1980).
[319] Wacker, W. E. C., Iida, C., Fuji,
K.: Nature 202: 659-662
(1964).
[320] Wacker, W. E. C., Iida, C., Fuwa,
K.: Nature 206: 90
(1965).
[321] Wacker, W. E. C., Parisi, A.
F.: New Engl. J. of Med.
278: 658-663, 712-717, 772-776 (1968).
[322] Wacker, W. E. C., Vallee, N.
L,: New Engl. J. Med. 259:
431-438, 475-482 (1958).
[323] Walker, A. R. P., Fox, F. W., Irving, J.
T.: Biochem, J., 42:
452-462 (1948).
[324] Walker, M. A., Page, L.: J.
Am. Diet. Assoc. 70: 260-266 (1977).
[325] Wallach, S., Carter, A. C.:
Am. J. Physiol. 200: 359-361 (1961).
[326] Walser, M.: Rev. Physiol.,
Biochem., Exp. Pharmacol. 59: 185-296
(1967).
[327] Wang, C. C. C., Kaucher, M., Wing,
M.: Am. J. Dis. Child. 52:
41-53 (1936).
[328] Wang, F. L., Wang, R., Khairallah, E. A., Schwartz,
R.: J. Nutr. 101:
1201-1202 (1971).
[329] Watchorn, E., McCance, R. A.:
Biochem. J. 26: 54-64 (1932).
[330] White, H. S.: J. Am. Diet.
Assoc. 55: 38-43 (1969).
[331] Widdowson, E. M.: Lancet
2: 1099-1105 (1965).
[332] — : J. Roy. Coll. Phys.
3: 285-298 (1969).
[333] Widdowson, E. M., Dickerson, J. W.
T.: In Comar, C. L. & Bronner,
F. (eds.). Mineral Metabolism 2, Academic Press, p. 2-247
(1961).
[334] Widdowson, E. M., McCance, R. A., Spray, C.
M.: Clin. Sci. 10: 113-125
(1951).
[335] Wilkinson, A. W., Harris, I.:
Brit. J. Surg. 56: 628 (1969).
[336] Workshop on Magnesium Intervention Trials. WHO
Collaborating Centre for Ref. on Studies of Cardiovascular
Disease in Relation to Water Quality. Ottawa, Canada. March 9-10
(1978).
[337] Young, V. R., Scrimshaw N. S.:
Am. J. Clin. Nutr. 32: 486-500 (1979).
[338] Zieve, L.: Ann N. Y. Acad.
Sci. 162: 732-743 (1969).
[339] — : Yale J. Biol. Med.
48: 229-237 (1975).
[340] Zieve, L., Doizaki, W. M., Stenroos, L.
E.: J. Lab. Clin. Med. 72:
261-267, 268-277 (1968).
[341] Zieve, F. J., Freude, K A., Zieve,
L.: J. Nutr. 107:
2178-2188 (1977).
[342] Zuckerman, L., Marquardt, G.
H.: Proc. Soc. Exper. Biol. Med.
112: 609-610 (1963).
[343] Zuspan, E.: J. Reprod. Med.
2: 116-139 (1969).
[344] Zuspan, F. P.: Clin. Obst.
Gyn. 9: 954-972 (1966).
[345] Zuspan, F. P., Pritchard, J. A., Quilligan, E.
J.: Contemp. Obst. Gyn. 6:
12-15 (1975).
Go to figures for this article, Magnesium Requirements in Human
Nutrition(figures)
This page was first uploaded to The Magnesium Web Site on
January 13, 2005
http://www.mgwater.com/

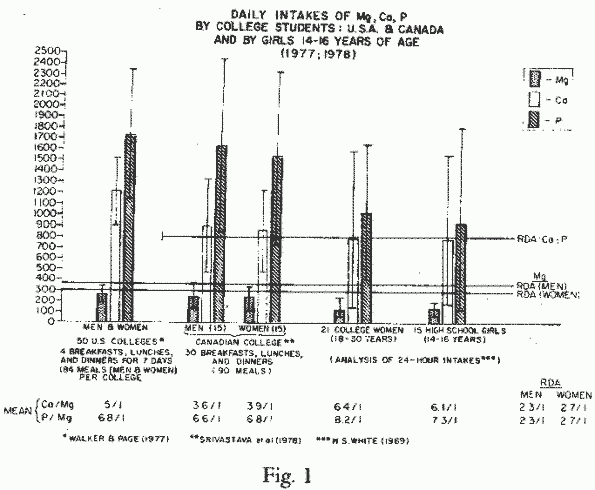{kind=link}
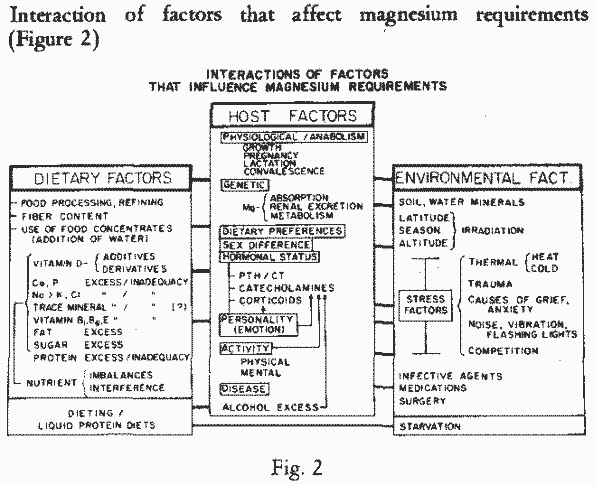{kind=link}
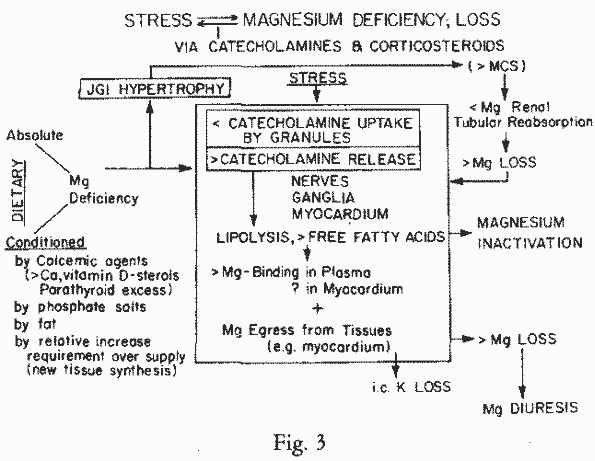{kind=link}
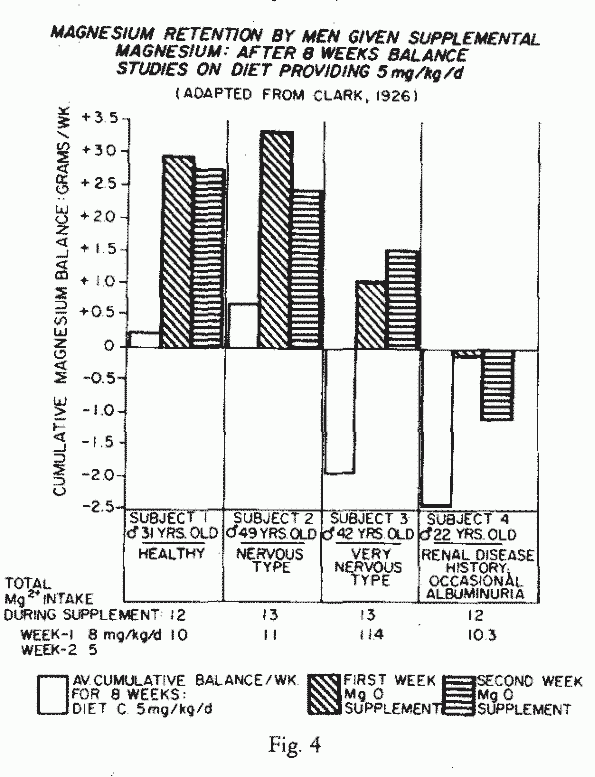{kind=link}
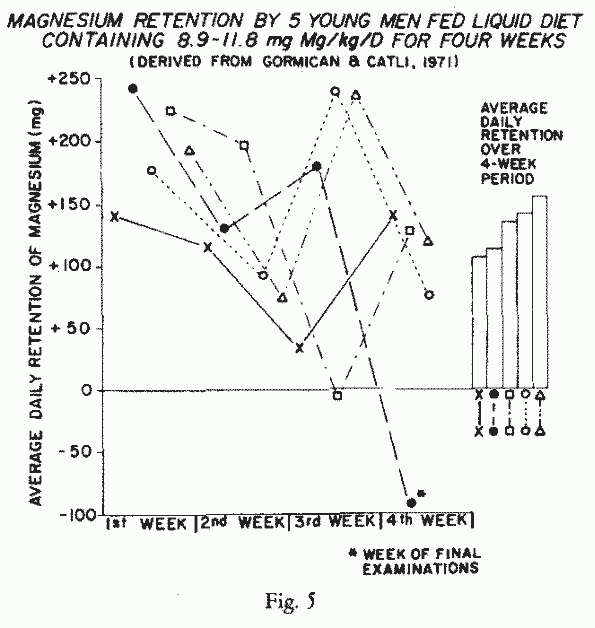{kind=link}
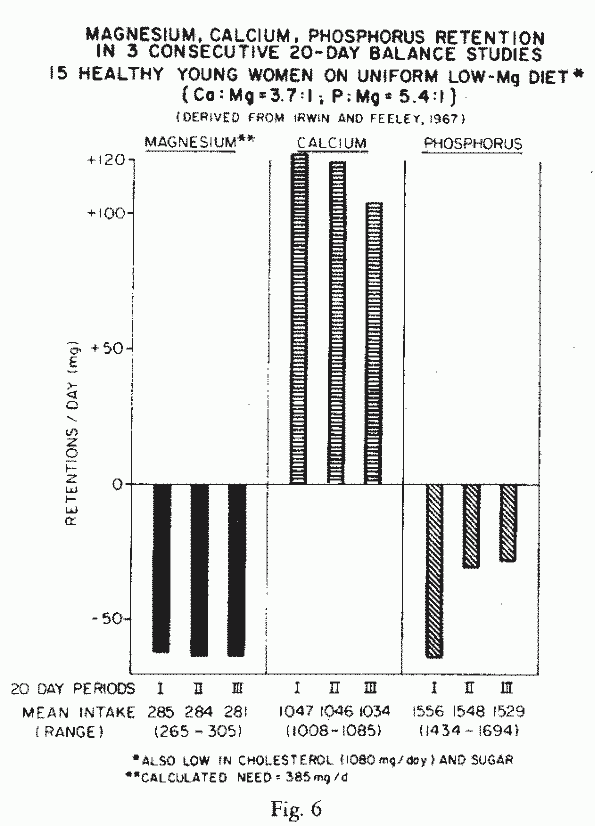{kind=link}
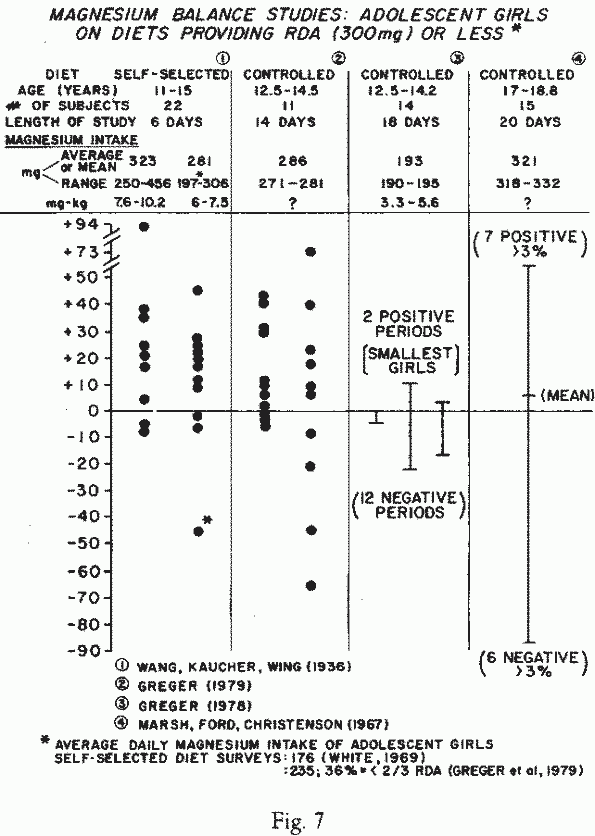{kind=link}
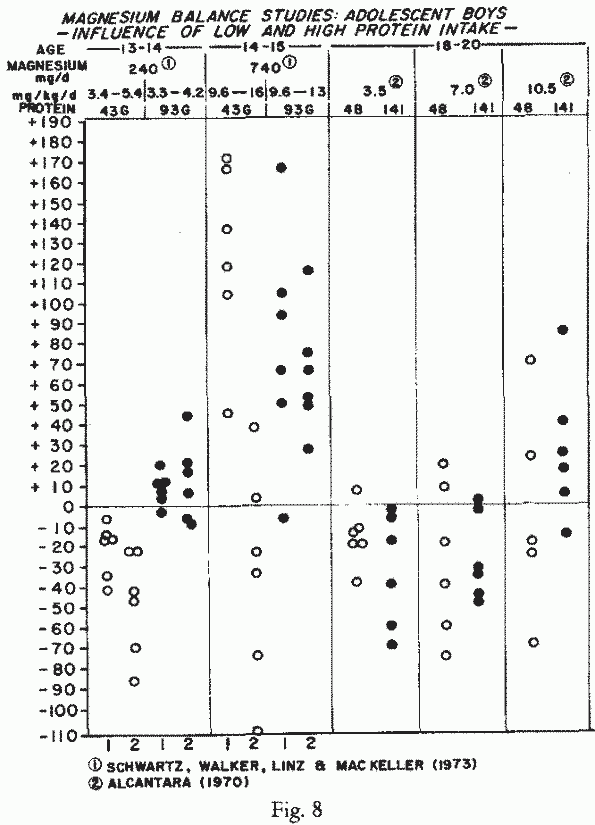{kind=link}
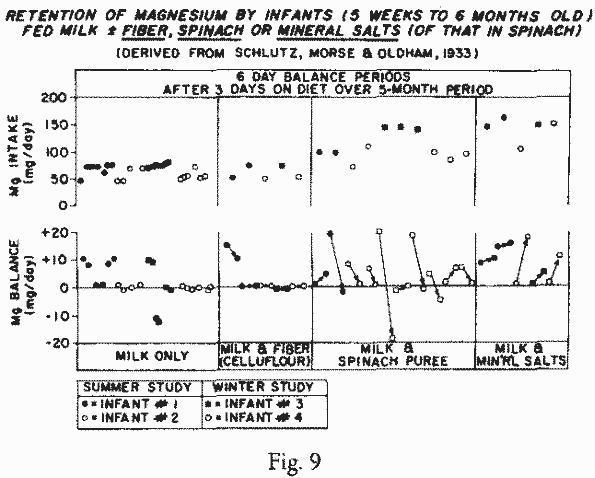{kind=link}
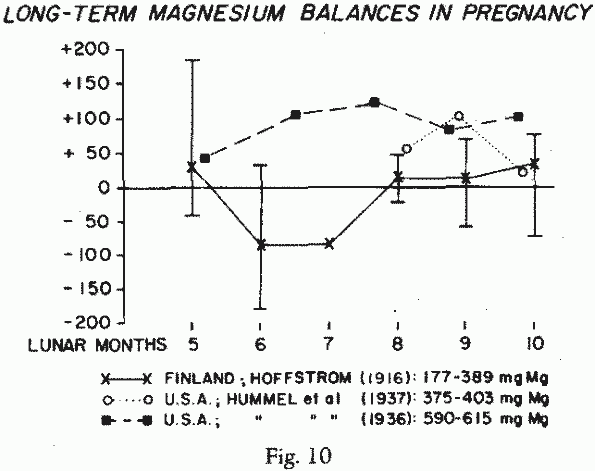{kind=link}
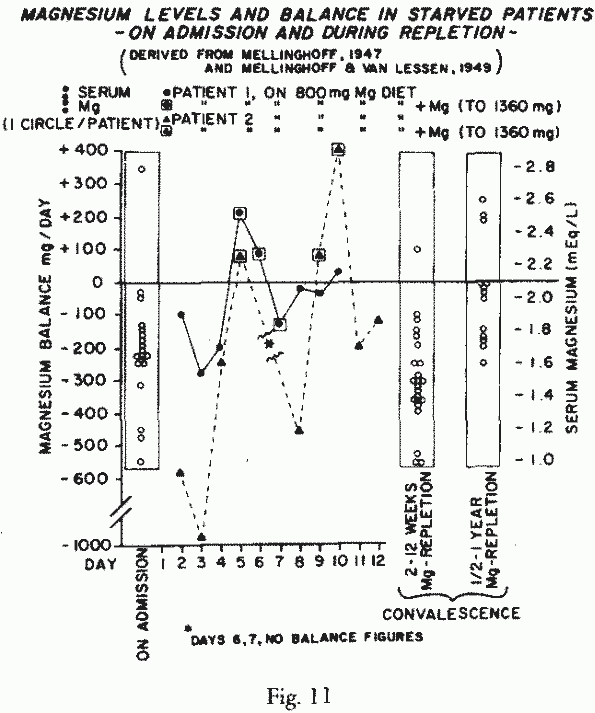{kind=link}
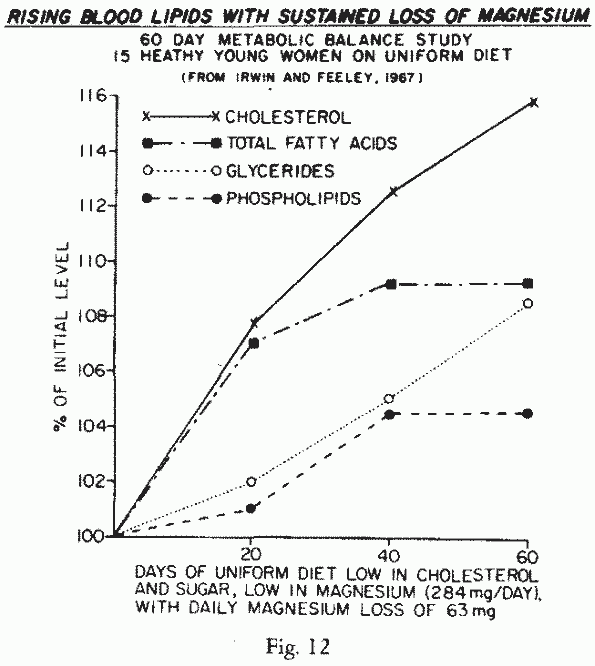{kind=link}
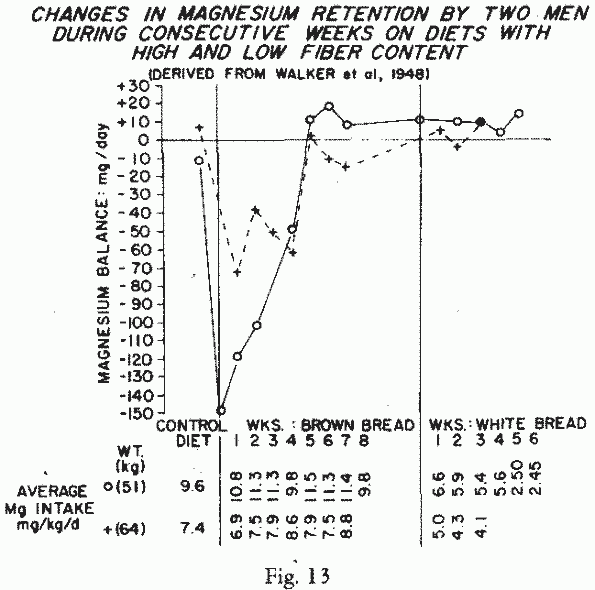{kind=link}
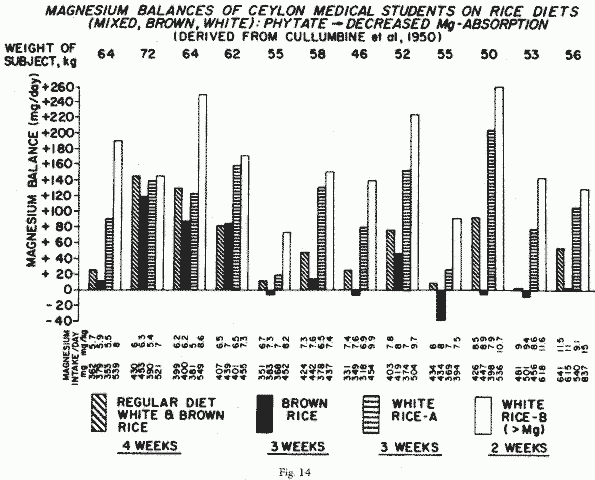{kind=link}
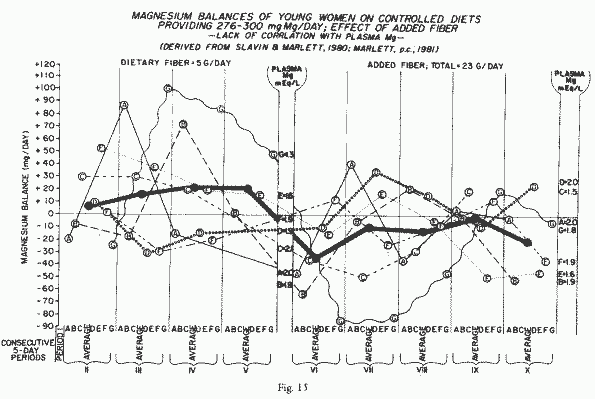{kind=link}